Inleiding tot het geslacht Taraxacum in Nederland
|
|
|
- Stijn Dijkstra
- 5 jaren geleden
- Aantal bezoeken:
Transcriptie
1 Inleiding tot het geslacht Taraxacum in Nederland door A.A. Sterk (Vakgroep Bijzondere Plantkunde, Universiteit van Amsterdam) Particularly in those genera that combine polyploidy with asexual reproduction such as Hieracium, Taraxacum, Rubus, Potentilla and Poa, the nightmare for the taxonomist will never be removed and species boundarieswill always be a matter of personal judgement. Stebbins, Inleiding Het geslacht Taraxacum vormt samen met enkele andere genera en soorten, waarvan de belangrijkste zijn Hieracium (havikskruid), Rubus (braam), Alchemilla vulgaris (vrouwenmantel), Poa pratensis (veldbeemdgras) en Ranunculus auricomus (gulden boterbloem) een zeer speciale groep van taxa binnen de Nederlandse flora. Al deze groepen vertonen een afwijkende voortplanting, die apomixis of agamospermie wordt genoemd. Hierbij worden in op het oog geheel normale bloemen zaden gevormd op asexuele wijze, d.w.z. zonder reductiedeling en bevruchting. Agamospermie is een voortplantingssysteem, dat groteinvloed heeft op de evolutie, de systematiek, de oecologie en de geografie van de betrokken groepen. Wat systematisch opvalt is het grote aantal taxa, meestal microspecies genoemd, die worden onderscheiden en de geringe verschillen tussen deze taxa. Vaak zijn de microspecies alleen na zorgvuldige bestudering en vergelijking met herbariummateriaal bevredigend op naam te brengen en vaak is dan nog hulp van specialisten onontbeerlijk. In algemene florawerken, ook de Nederlandse, worden de microspecies meestal niet alle opgenomen; vaak worden alleen de goed herkenbare en ruim verspreide soorten genoemd ofwordt volstaan met de aanduiding van soortsgroepen of secties (Flora Europaea). Het is daarom verheugend dat in de Flora Neerlandica de tot nu toe beschreven microspecies van Taraxacum worden opgenomen, waardoor gedetailleerd onderzoek over het geslacht mogelijk wordt en Nederland betrokken kan worden in internationale studies over de systematiek, de oecologie en de plantengeografie van Taraxacum. 53
2 2. De classificatie in historisch perspectief Lange tijd is gedacht dat het geslacht Taraxacum soortenarm en eenvormig was. Linnaeus (1753) onderscheidde Taraxacum niet als apart geslacht, bracht de maar paardebloemen onder bij Leontodon, als de soort Leontodon taraxacum. De Candolle (1838) gaf 85 jaar later een bewerking van het geslacht in 3 secties met 30 soorten en deze indeling bleef gezaghebbend tot de laatste eeuwwisseling. Het was ook het uitgangspunt voor Handel-Mazzetti (1907) voor zijn Monographie der Gattung Taraxacum, waarin hij het geslacht in 11 secties en 57 soorten verdeelt. Dit waren z.g. grote oflinneaanse soorten. De voor grote Taraxacum ontstond evenwel belangstelling tijdens de laatste eeuwwisseling, toen in Zweden en Denemarken door Ostenfeld en Raunkiaerwerd aangetoond dat paardebloemen zich op bijzondere wijze voortplanten. Ostenfeld vond paardebloemen zonder pollenkorrels, die toch zaden voortbrachten, terwijl Raunkiaeraantoondedatook gecastreerde bloemenzaden kunnen vormen. Beide onderzoekers kwamen tot de conclusie dat de zaden door agamospermie zouden worden gevormd, d.w.z. op een asexuele wijze zonder bevruchting; spoedig werd dit ook door cytologisch onderzoek, o.m. door Murbeck in 1904, bewezen. Het onderzoek bij Taraxacum werd nu snel geïntensiveerd en in de taxonomie ging men over tot het onderscheiden van de eng begrensde kleine soorten, waarvan het aantal snel steeg tot in de honderden. Dit onderzoek vond vooral plaats in de Scandinavische landen en Finland. Buiten Noordwest-Europa gebruikte men overwegend nog de grote soorten van Handel-Mazzetti. Dit veranderde toen prof. dr. ir. J. L. van Soest in dejaren dertig met zijn onderzoek aan Taraxacum begon. Hij sloot met zijn werk aan bij de Noordeuropese ging ook over tot het onderscheiden van kleine soorten. Van zijn scherp auteurs en inzicht in de vormvariatie bij Taraxacum en zijn uitgebreide publicistische arbeid is een grote invloed uitgegaan, als gevolg waarvan de kleine soorten in Europa en daarbuiten nu door velen worden aanvaard. Een uitzondering hierop vormen Amerikaanse auteurs, die nog vaak de grote soorten van Handel-Mazzetti gebruiken. Ook in de agrarische literatuurworden meestal grote soorten ofsecties gebruikt. Door de voortgang van het onderzoek in de laatste decennia is het aantal secties en soorten sterk toegenomen. Er wordennu23 secties onderscheidenmet soorten en Taraxacum behoorthiermeetot de grote composieten-geslachten (Dole, 1977). Het geslacht komt in alle werelddelen voor; in Zuid-Azië enin het grootste deel van Amerika en Afrika zijn paardebloemen evenwel niet inheems, maar door de mens ingevoerd en ingeburgerd. In Europa, het best onderzochte deel van het areaal, zijn ca soorten beschreven in 21 secties (Richards & Sell, 1976). Van 2 Europese secties is er een recente monografie, nl. van de sectie Palustriamet 48 soorten (Van Soest, 1965) en de sectie Erythrosperma met 115 soorten (Dole, 1973). Bij dit alles moet evenwel worden opgemerkt dat van grote delen van Europa nog geen overzicht van de Taraxacum-flora bestaat; van de volgende landen bestaan meer of minder uitvoerige Taraxacum-bewerkingen: Noorwegen, Zweden, Finland, Denemarken, Ijsland, Groot-Brittannië, Nederland, België, Polen, Rusland, en Zwitserland (zie Richards & Sell, l.c.). Gezien het bovenstaande valt het dan ook te verwachten dat het aantal soorten sterk zal nog stijgen bij toekomstig onderzoek. 54
3 Omdat veel microspecies recent pas zijn onderscheiden en nog slechts een klein deel van het areaal van Taraxacum relatief goed is onderzocht, kunnen weinig plantengeografische conclusies worden getrokken met betrekking tot de microspecies. Duidelijk is wel dat er een grote variatie bestaat in areaalgrootte. Sommige soorten hebben een areaal dat zich over grote delen van Europa uitstrekt, b.v. T. rubicundum, die voorkomt van Zuid-Scandinaviëtot in het Middellandse Zeegebied en die ingeburgerd is in delen van de U.S.A. Andere soorten hebben een klein areaal, zoalst. agaurum, dieendemisch is in Nederland en die alleen wordt gevonden in het Duindistrict van Noordwijk tot Voorne. Er zijn zelfs soorten bekend, die slechts op één plaats zijn gevonden, zoals T. westhoffii op Terschelling. Ook van de oecologie der meeste microspecies weten we betrekkelijk weinig. Bekend is wel dat Taraxacum-soorten in zeer uiteenlopende habitats kunnen voorkomen, zowel in het laagland als in het hooggebergte, in droge en vochtige, voedselrijke en voedselarme habitats, onder oceanische, continentale, arctische en mediterrane omstandigheden. Duidelijk is dat vele soorten gebonden zijn aan anthropogene habitats. Hierbij is het opmerkelijk datin één graslandperceel 20 ofmeer soorten van de sectie V ulgaria kunnen voorkomen en dit gegeven plaatst de oecologen voor het probleem van de coëxistentie van zeer nauw verwante soorten in één biotoop. Er zijn evenwel ook soorten, die weinig anthropogene invloeden kunnen verdragen en die b.v. in ons land tot de zeldzame en bedreigde soorten behoren, zoals Palustria- en Spectabilia-soorten van schraalgraslandenvan de Pijpestrootjes-orde (Molinietalia). Een goed overzicht van de huidige kennis van het geslacht Taraxacum geven Doll (1974, 1977), Richards & Sell (1976) en Richards (1973). Voor de begingeschiedenis van de Taraxacum-classificatiein ons land moge worden verwezen naar Van Soest (1939, 1942 en 1955). Bij Vuyck (1902) vinden we een zeer summiere en naar huidige maatstaven zeer onvolledige indeling. Hetzelfde geldt voor de oudere drukken van de flora s van Heukels en van Heukels-Wachter. In Heukels- Wachter (1934) wordt één Taraxacum-soort onderscheiden, Web. met 3 ondersoorten, t.w. subsp. palustre (overeenkomend Taraxacum officinale met de sectie Palustria en sectie Spectabilia?), subsp. officinale (overeenkomend met de sectie Vulgaria) en subsp. levigatum (overeenkomend met de secties Obliqua en Erythrosperma). Een moderne indeling van de Nederlandse Taraxaca met een verkorte sleutel van Dahlstedt geeft Van Soest (1939). Deze indeling noemt de Vulgaria, Palustria, Obliqua, Dissimilia en Erythrosperma; de Spectabilia zijn er nog niet bij. Deze groep verschijnt in Van Soest (1942) enalle in Nederlandvoorkomendesecties zijn dan genoemd. De Dissimilia worden later ondergebracht bij de Erythrosperma. In 1942 zijn in totaal voor Nederland 21 soorten beschreven, waaronder 6 Vulgaria-soorten; in 1957 is dit opgelopen tot 42 soorten, waaronder 16 Vulgaria-soorten. Voor de verdere ontwikkeling van de Taraxacum-systematiek is de samenwerking tussen prof. Van Soest en de herena. Hagendijk en H. A. Zevenbergen van veel belang. Deze samenwerking dateertvan het laatste deel van de jaren zestig. In 1975 publiceerden Hagendijk, Van Soest & Zevenbergen in Flora Neerlandica 42 soorten en wel 1 soort van de sectie Obliqua, 22 van de sectie Erythrosperma, 10 van de sectie Palustria en 9 soorten van de sectie Spectabilia; voor verdere publicaties van Taraxacum-soorten, 55

4 zie Vogelenzang (1978). In het hierna volgende laatste deel van Taraxacum, dat de sectie Vulgaria omvat, worden 153 soorten genoemd. Er zijn nu in totaal voor Nederland 196 soorten bekend in 5 secties; hiervan zijn door Van Soest 16 soorten beschreven, door Van Soest met een buitenlandse auteur 1 soort, door Hagendijk, Van Soest en Zevenbergen 58 soorten, doornederlandse auteurs, exclusiefvan Soest, 2 soorten en dooruitsluitendbuitenlandse auteurs 119 soorten. Het blijkt dusdat het grootste deel van dein land voorkomen- ons de soorten, nl. 119, doorandere dan Nederlandse auteurs zijn beschreven; deze soorten komen ook buiten ons land voor. Maar omgekeerd zijn er voor Nederland soorten beschreven, die ook in het buitenlandworden gevonden. 3. De voortplanting Zoals reeds is opgemerkt vinden we binnen Taraxacum een bijzondere wijze van voortplanting, nl. agamospermie. Van agamospermie bestaan verschillende vormen en de vorm die bij Taraxacum voorkomt heet diplospore parthenogenese. De diplospore parthenogenese zal hieronder kort worden besproken en zal worden vergeleken met de normale sexuele voortplanting (voor details, zie Rutishauser, 1967). a. De sexuele voortplanting In het vruchtbeginsel van Taraxacum bevindt zich één zaadknop, die bestaat uit een nucellus met daaromheen twee integumenten die de z.g. micropyle openlaten. In de nucellus ontstaat de embryozakmoedercel (EMC) en deze diploïde (2n) EMC maakt een reductiedeling (meiosis) door. Bij de eerste reductiedeling paren de homologe chromosomen, waarna er van ieder paar homologe chromosomen één in de dochterkern terecht komt. Hierdoor zijn er na afloop de deling van twee haploïde (n) dochterkernen ontstaan (fig. 49). Daarna volgt de tweede meiotische deling, die het karakter heeft van een normale celdeling en er ontstaan dan 4 haploïde embryozakcellen. Van deze 4 cellen gaan er 3 te gronde en in de overblijvende cel treedt driemaaleen kerndeling op. Er wordt zo een 8-kernige embryozak gevormd, waarin de haploïde kernen op een speciale wijze zijn gerangschikt (zie fig. 49). Aan de zijde van de micropyle vinden we de eicel met de twee synergiden, aan de andere kant van de embryozak de 3 antipoden en in het centrum de 2 poolkernen. Kort voor de bevruchting fuseren de 2 poolkernen tot de secundaire kiemzakkern. In de helmknoppen van de meeldraden ontstaan Fig. 49. Schema van de sexuele voortplanting en de diplospore parthenogenese bij Taraxacum. 56
5 uit de pollenmoedercellen na reductiedeling vier haploïde pollenkorrels. De pollenkorrels groeien, na bestuiving, op de stempel uit tot pollenbuizen. Hierin bevinden zich één vegetatieve cel en twee zaadcellen. Een van de zaadcellen (n) fuseert met de eicel (n) tot een zygote, die nu diploïd (2n) is en de andere (n) fuseert met de secundaire kiemzakkern (2n), die nu triploïd (3n) is. Uit de zygote ontwikkelt zich de nieuwe plant en uit de bevruchte secundaire kiemzakkern ontwikkelt zich het endosperm, dat het reservevoedsel voor het embryo is. We noemendit de z.g. dubbele bevruchting. b. De agamospermie Bij de eerste deling van de EMC paren de homologe chromosomen niet; zij worden niet over twee dochterkernen verdeeld, maar komen in één z.g. restitutiekern terecht, die diploïd (2n)is (zie fig. 49). De tweede meiotische deling verloopt normaal en er worden zo twee diploïde (2n) embryozakcellen gevormd. Dit verschijnsel heet diplosporie. Eén van de twee embryozakcellen deelt verder en er wordt weer een 8-kernige embryozak gevormd, nu niet met haploïde kernen, maar met diploïde. De eicel groeit nu zelfstandig, d.w.z. zonder bevruchting, uit tot een nieuwe diploïde plant. De twee poolkernen fuseren weer tot de secundaire kiemzakkern, die nu tetraploïd (4n 2n + 2n) is; uit deze kiemzakkern groeit zelfstandig, zonder bevruchting, het endosperm dat het reservevoedsel voor de kiemplant is. Het zelfstandig uitgroeien van de eicel, dus zonder bevruchting, noemen we parthenogenese (parthenos maagd). Bij agamosperme paardebloemen is, zoals duidelijk zal zijn, bestuiving dus niet noodzakelijk. De beschreven vorm van agamospermie heet diplospore parthenogenese. Om vergelijking met het schema van de sexuele voortplanting mogelijk te maken is bij de bespreking van de agamospermie ook steeds gesproken van haploïd en diploïd. Dit zou verwarring kunnen wekken, omdat agamosperme paardebloemen steeds polyploïd zijn en wel meestal triploïd (zie verder onder Chromosoomgetallen). Daarom ter verduidelijking het volgende. Het is gebruikelijk om ook bij polyploïde chromosoomgetallen de begrippen haploïd en diploïd te gebruiken, aangegeven met resp. n en 2n; 2n geeft dan het aantal chromosomen aan dat in de lichaamscellen aanwezig is. Daarnaast wordt bij polyploïde reeksen het begrip basisgetal gebruikt, aangeduid door x. Vaak vinden we dat de chromosoomgetallen van een polyploïde reeks veelvouden zijn van het basisgetal. Het basisgetal bij de paardebloem is x 8. Veel paardebloemen zijn triploïden, d.w.z. dat de homologe chromosomen in drievoud aanwezig zijn, de notitie voor een triploïde paardebloemensoort is: 2n 3x 24 (3 x 8). Nu komt er naast de besproken agamospermie, waarbij alle bloemen diplospoor zijn, bij paardebloemen ook een vorm voor, vermoedelijk op beperkte schaal, waarbij in één bloemhoofdje sommige EMC s een diplospore ontwikkeling doormaken en andere een normale meiosis hebben. Met een normale meiose wordt bedoeld dat de eerste reductiedeling ook werkelijk tot een kerndeling en een celdeling leidtevenals de tweede reductiedeling. Meestal is de eerste reductiedeling dan sterk gestoord, omdat bij de triploïde paardebloemen de homologe chromosomen in drievoud aanwezig zijn, waardoor paring chromosomen moeilijkheden geeft. van In de gestoorde reductiedelingen zien we dan ook dat de paring geheel verschillend uitvalt; er zijn ongepaarde chromosomen (univalenten), gepaarde chromosomen (bivalenten) en drie bijeenliggende chro- 57
6 mosomen (trivalenten). De verdeling van de chromosomen over de dochtercellen bij de eerste reductiedeling is als gevolg hiervan zeer onregelmatig, sommige cellen hebben te veel chromosomen, andere te weinig. Als resultaat hiervan vinden we ook bij de eicellen veel variatie in chromosoomgetal; zeer belangrijk is nu dat hierbij ook haploïde (n 8) getallen kunnen voorkomen. Deze haploïde eicellen blijken een volledig chromosoomset te kunnen hebben en gedragen zich dan als geheel normale eicellen. Zij kunnen bevrucht worden met haploïde zaadcellen en zo kunnen diploïde en sexuele paardebloemen ontstaan. Hierbij zijn dus uit triploïde agamosperme planten sexuele diploïde planten ontstaan. We noemen deze planten facultatief agamosperm; de eerder besproken planten met uitsluitend restitutionele reductiedelingen heten obligaat agamosperm. Bij facultatieve agamospermie is dus wel bestuiving nodig, nl. voor de sexuele bloemen. De facultatieve agamospermie heeft een belangrijke functie; zij kan immers zorgen voor nieuwe genetische variatie, waardoor de agamospermepopulaties toch mogelijkheden voor genetische aanpassing behouden. Opvallend bij Taraxacum is nu dat er bij veel agamosperme soorten grote verschillen bestaan tussen de meiosenin de helmknoppen en in de zaadknoppen. Bij de meerderheid der soorten vinden we in de zaadknoppen de bovenbeschreven restitutionele meiosen, waarbij dus eenrestitutiekernwordt gevormd (diplosporie). Bij een minderheid daarentegen komen ook normale reductiedelingen, z.g. reductionele meiosen, voor; deze reductiedelingen zijn dan meestal sterk gestoord. In de meeldraden is het omgekeerde het geval: hier komen restitutionelemeiosen weinig voor en zijn gestoorde onregelmatige reductionelemeiosen algemeen. Hierbij treedt dan in meerdereofmindere mate paring van de homologe chromosomen op met als resultaat dat de verdeling van de chromosomen over de dochterkernen bij de eerste reductiedeling onregelmatig is. Er ontstaan dan niet alleen normale pollenkorrels, ook pollenkorrels maar met te weinig chromosomen en pollenkorrels met teveel chromosomen. De pollenkorrels met te weinig chromosomen zijn kleiner dan normaal en pollenkorrels met teveel chromosomen zijn groter dan normaal. Is de meiosis normaal en ongestoord, zoals bij diploïden, dan varieert de diameter van het pollen meestal van 30 fim met een gemiddelde, dat ligt bij ongeveer 35 Hm; is de meiosis gestoord, zoals dat bij triploïden het geval is, dan varieert de diameter van 1570 /tm met een gemiddelde, dat ligt tussen 35 /im of /tm. Uit de variatie in pollengrootte kunnen we afleiden of we te maken hebben met diploïde sexuele planten, dan wel met agamosperme, die meestal triploïd zijn. Overigens bestaan er gradaties in regelmatigheid van het pollen. Volgens Richards (1972) is het pollen van triploïd facultatief agamosperme planten in variatiebreedte en gemiddelde moeilijk te onderscheiden van datvan diploïd sexuele planten. Zelden vindenwe een gemiddelde dat hoog ligt en wel bij 45 fim; hier zou sprake kunnen zijn van agamosperme tetraploïden. Op basis van pollenonderzoek is het met een hoge mate van betrouwbaarheid mogelijk gebleken om in de natuur en aan herbariummateriaal (bloemknoppen) vast te stellen of een individu sexueel dan wel agamosperm is (voor details, zie Den Nijs & Sterk, 1980). Ook voor een groot aantal Nederlandse Taraxacum-soorten is op het Hugo de Vries-laboratorium (Amsterdam) de sexualiteit/agamospermie via variatie in pollengrootte aan herbariumplanten bestudeerd (Rijksherbarium, Leiden, en Herba- 58
7 rium Van Soest, opgenomen in het Rijksherbarium). De resultaten hiervan worden in de hierna volgende systematische bewerking van de sectie Vulgaria bij de soorten vermeld. Verreweg de meeste onderzochte soorten hebben duidelijk onregelmatig pollen (code PO) en zijn zeer waarschijnlijk triploïd agamosperm. Er zijn evenwel enkele planten gevonden met zeer regelmatig pollen (PR) met een variatiebreedteen een gemiddelde, die gelijk zijn aan die van sexuele planten. Het betreft de volgende soorten: T. alatum van de St. Pietersberg, T. angustisquameum van Epen, T. croceiflorum van Ommen en van Eist en T. limburgense van Mechelen. Van T. limburgense zijn bij Mechelen, langs de Geul, levende planten verzameld, die2n 16 chromosomen bleken te hebben; deze planten zijn dus diploïd en sexueel. Het betreft hier waarschijnlijk dezelfde vindplaats als die waar ook het onderzochte herbariummateriaalis verzameld. Uit deze waarneming wordt het zeer waarschijnlijk, dat ook de andere planten met regelmatig pollen, die behoren tot de bovengenoemde soorten, diploïd en sexueel zijn. Aangezien de herbariumplanten een relatiefkleine steekproef vormen van de Nederlandse paardebloemen, kan verwacht worden dat meer soorten in ons landsexuele individuen zullen hebben. Van de soorten T. alatum en T. croceiflorum zijn óók planten met onregelmatige pollen gevonden; deze planten zijn duidelijk triploïd. Het voorkomen van diploïde èn triploïde planten binnen één soort kan wijzen facultatieve op agamospermie. In Nederland zijn tenslotte ook planten aangetroffen met een pollenbeeld dat veel overeenkomst met dat van de sexuele planten vertoont. Volgens Richards (lx.) zouden deze planten facultatief agamosperm kunnen zijn. In afwachting van verderonderzoek zijn deze planten gerekend tot de groep met onregelmatig pollen. Bij obligate agamospermie is bestuiving niet nodig en het pollen is dan ook functieloos geworden. Bij een aantal soorten vinden we als gevolg hiervan in de helmknoppen een volledige degeneratie van de meiose met als resultaat dat er in het geheel geen pollen meer wordt gevormd. Dit doet zich ook bij Nederlandse Taraxacum-soorten voor en het al ofniet aanwezig zijn van pollen is in de determinatietabellendan ook een belangrijk diagnostisch kenmerk. De sexuele planten Taraxacum hebben protandrische bloemen, die zelfincompatibel zijn. Bij protandrische bloemen komt het pollen vrij vóórdat de stempel ontvanke- van lijk is, terwijl bij bloemen die zelfmcompatibel zijn zelfbestuiving niet tot zaadzetting leidt. Zowel protandrie als zelfmcompatibiliteit hebben de functie om kruisbestuiving te verzekeren. De bloemen van Taraxacum hebben een bestuivingsmechanisme, dat bij veel composieten voorkomt. Hierbij zijn de helmknoppen van de meeldraden zijdelings vergroeid tot een z.g. antherenkoker. De twee samengevouwen stempellobben zitten onderin de koker zoals een zuiger in een cilinder. Het pollen wordt boven de zuiger in de cilinder uitgestort. Bij de verdere ontwikkeling van de bloem gaan de stijl en de stempel groeien en duwen zo het pollen uit de antherenkoker. De stijl draagt haartjes; deze haartjes vindenwe ook op de buitenzijde, dit is de niet-ontvankelijke kant van de stempellobben. Met deze veeghaartjes wordt het pollen uit de koker geveegd, waarbij het pollen tussen de haartjes blijft vastzitten. Als de stempellobben geheel uit de koker zijn gegroeid wijken uit ze elkaar, waardoorhet ontvankelijke deel van de en stempel vrijkomt bestoven kan worden met vreemd pollen. De stempellobben groeien verder uit en krullen naar achteren om (fig. 50), zodat tenslotte zelfbestuiving op kan treden. Deze zelfbestuiving 59

8 24), 32) 16) Fig. 50. Schema van de ontwikkeling van een bloempje bij Taraxacum (zie tekst) leidt evenwel bij Taraxacum, tengevolge van zelfincompatibiliteit, niet tot zelfbevruchting. In een hoofdje begint de bloei bij de randbloemen en geleidelijk komen daarna de meer centrale bloemenin bloei. In één hoofdje zijn de verschillende ontwikkelingsstadia de van afzonderlijke bloemen meestal goed te zien. Het gehele bestuivingsmechanisme, zoals dathierboven beschreven is voor de sexuele planten, is bij de agamosperme planten Taraxacum geheel intact gebleven. De van agamosperme planten vormen daarbij meestal veel pollen en nectar en zijn in het voorjaar, als er nog weinig bloeiende plantesoorten zijn, een rijke voedingsbron voor allerlei insecten. Paardebloemenkunnen zelfs optreden als concurrenten bij de bestuiving van fruitbomen (appels) en worden danook in boomgaarden wel bestreden (Faegri & Van der Pijl, 1979). 4. Chromosoomgetallen Bij Taraxacum bestaat er een strenge correlatie tussen de wijze van voortplanting en de ploïdiegraad. Taraxacum is een zg. polyploïd complex met als basisgetal x 8. We vinden in het complex diploïde (2x soorten en verder een reeks soorten met chromosoomgetallen lopend van 3x 24 tot lox 80. De meeste soorten zijn triploïd (3x tetraploïden (4x komen eveneens vrij veel voor, maar de hogere ploïdiegraden zijn, evenals de diploïden, zeldzaam. Tot nu toe is steeds gevonden dat de diploïde soortensexueel zijn en de polyploïde agamosperm, obligaat offacultatief. Zeer waarschijnlijk zijn de polyploïden afgeleid van diploïde sexuele via hybridisatie en polyploïdisatie. In Hagendijk, Van Soest & Zevenbergen (1975) worden bij veel soorten van de secties Erythrosperma, Obliqua, Palustria en Spectabilia chromosoomgetallen vermeld. De meeste tellingen zijn evenwel verricht aan buitenlandse planten; enkele aan Nederlandse (Hou-Ltu, 1963). Latere tellingen aan Nederlandse planten zijn verricht door Bosch (1979), Den Nijs, Sterk & Van der Hammen (1978), Hagendijk, Oosterveld & Zevenbergen (1978), Fengler (1978) en Mooren (1981). Voor Nederland zijn nu van de secties Erythrosperma (E), Obliqua (O), Palustria (P) en Spectabilia (S) de volgende chromosoomgetallen bekend: T. anglicum (S) 2n 24; T. brachyglossum 60
9 24; 32) 24) 24) 23) 24) 24) 48) (E) 2n 24, 32 (33, 34); T. commixtum (E) 2n 24, 32 (64); T. dunense (E) 2n 24; T. frisicum (P) 2n 24; T. hollandicum (P) 2n 24(25, 26),32;T. hygrophilum (S) 2n 24, 40; T. lacistophyllum (E) 2n 24 (23, 25, 26); T. laetiforme (E) 2n T. maculigerum (S) 2n 24; T. maritimum (P) 2n 24 (25); T. nordstedtii (S) 2n 48 (46, 47, 49, 50); T. oxoniense (E) 2n 24; T. palustre (P) 2n 24 (23, 25), 32 (31, 64); T. proximum (E) 2n 24; T. rubicundum (E) 2n 24 (26); T. scanicum (E) 2n 24; T. silesiacum (E) 2n 24 (23, 26), 32; T. taeniatum (E) 2n 24 (25); T. tortilobum (E) 2n 24 (25); T. vansoestii (E) 2n 24, 32. Van de sectie Vulgaria zijn chromosomen voor Nederland geteld doorbosch (1979), Fengler (1978), Fengler & Sterk (1978) en Mooren (1981). De getallen worden in het hiernavolgende deel bij de Vulgaria-soorten vermeld. Wanneerwe de gegevens uit Nederland samenvatten, dan blijkt het volgende: 1. Sectie Obliqua: 1 soort, alleen triploïden (2n gevonden. 2. Sectie Erythrosperma: 23 soorten, waarvan 13 onderzocht, hiervan 9 soorten triploïd (2n 24) en 4 soorten zowel triploïd (2n als tetraploïd (2n 32). 3. Sectie Palustria: 10 soorten, waarvan 4 onderzocht; hiervan 2 soorten triploïd (2n 24) en 2 soorten zowel triploïd (2n 24) als tetraploïd (2n 32). 4. Sectie Spectabilia: 9 soorten, waarvan 4 onderzocht; hiervan 2 soorten triploïd (2n 24), 1 soort triploïd (2n en pentaploïd (2n 40) en 1 soort hexaploïd (2n 48). 5. Sectie Vulgaria: 153 soorten, waarvan 55 onderzocht; hiervan 54 soorten triploïd (2n en 1 soort diploïd (2n 16). Bij alle soorten, met uitzondering van T. nordstedtii (2n en T. limburgense (2n 16), komen triploïde individuen voor. Er zijn geen morfologische verschillen aangetoond tussen de triploïden en de tetraploïden binnen één soort. Bij geen enkele soort zijn er uitsluitend tetraploïden gevonden. Bij veel soorten komen ook aneuploïde getallen die met voor, één ofenkele chromosomen naar boven of naarbeneden het van triploïde (2n 24) of tetraploïde getal (2n afwijken, b.v. 2n 23 of 2n 25. Dit kan belangrijke consequenties hebben, omdat verlies van één chromosoom kan leiden tot herstel van de sexualiteit, zoals is aangetoond door Sorensen & Gudjonsson (1946) enkele voor Vulgaria-soorten. Bij deze hypotriploïden trad weer reductiedeling op, waarbij haploïde eicellen ontstonden, die door haploïd pollen werden bevrucht. Hieruit kwamen diploïde sexuele planten voort. Op het chromosoom dat verloren ging lagen kennelijk de reductiedeling-onderdrukkende genen; hierdoor kregen de normale meiosisgenen in deze hypotriploïden weer de overhand. Deze hypotriploïden konden door Sorensen & Gudjonsson morfologisch onderscheiden worden van de triploïden en Hagendijk, Van Soest & Zevenbergen sluiten niet uit, dat deze hypotriploïden ook in Nederland voorkomen; zie hiervoor het navolgende deel onder T. dahlstedtiien T. polyodon. Wanneer dit zo is, dan zou ook deze sexualiteitmet de mogelijkheid van hybridisatie belangrijke consequenties kunnen hebben voor het ontstaan van nieuwe taxa in ons land. We zien hier het verschijnsel dat de reductiedeling bij een hypotriploïd (2n zodanig kan verlopen, dat hieruit functionele eicellen en zaadcellen ontstaan, die n 8 ofn 16 chromosomen hebben, ofeen aantal dat er dichtbij ligt. De eicellenen zaadcellen met tussenliggende chromosoomgetallen zijn meestal abortief. We vinden dit ook bij, zoals reeds is besproken, facul- 61
.")
10 tatief agamosperme triploïden. Wel moet worden opgemerkt dat de hypotriploïden, die Sorensen & Gudjonsson in hun proeftuin vonden, zeer zelden voorkwamen en dat de planten duidelijk verminderd vitaal waren (Sorensen, 1958). Het staat niet zonder meer vast dat deze planten, die onder omstandigheden van concurrentie in de natuur weinig kansen hebben, kunnen bijdragen tot de vorming van een nieuwe variabiliteit, eventueel van nieuwe taxa; onmogelijk is het evenwel niet. Wanneer wij de voortplanting en de ploïdiegraad van de sectie Vulgaria in West- Europa bezien, dan blijkt er merkwaardige verdeling in sexualiteit een en agamospermie en in de eraan gekoppelde ploïdiegraden te bestaan (Den Nijs & Sterk, 1980). We kunnen dat zeggen globaal langs de lijn Nantes - - ten zuiden van Parijs Stuttgart - Passau een overgangszone bestaat, die twee gebieden scheidt. Ten zuiden van deze zone vinden we diploïde sexuele populaties en gemengd diploïd/sexuele-triploïd/agamosperme populaties en ten noorden ervan triploïd/agamosperme populaties met slechts incidenteel diploïden (Engeland en Zweden, zie Richards, 1972 en Nederland). Nederland valt dus in het voornamelijk triploïd/agamosperme gebied. Een verklaring voor deze plantengeografische verdeling van voortplantingssystemen en ploïdiegraden is er nog niet. 5. De fenotypische plasticiteit Alle soorten, maar in het bijzonder die van de sectie Vulgaria, vertonen een grote fenotypische plasticiteit, met name in het blad. De variatiein bladvorm kan verschillende oorzaken hebben. Er is variatie samenhangend met de leeftijd van de met plant, het seizoen en met het milieu. Juveniele planten hebben meestal weinig ingesneden bladeren (fig. 51a). Sommige soorten in graslanden houden in het eerste levensjaar, waarin ze vaak niet bloeien, deze praktisch oningesneden bladeren. Volwassen planten hebben in het voorjaar tijdens de bloei karakteristiek ingesneden bladeren (fig. 5IA). Al het eind aan van de bloei (meestal half mei-eind mei) sterven deze bladeren en er verschijnen nieuwe spruiten. Deze spruiten vormen eerst niet- of weinig ingesneden bladeren (fig. 5 ld), de later gevormde bladeren worden steeds meer ingesneden (fig. 51c) tot tenslotte weer normaal ingesneden bladeren zijn gevormd (fig. 51 b) Na de. zomer gaat de vorming van ingesneden bladeren bij veel soorten door. Ook in de winter zijn de bladeren ingesneden, veelal is dan de bladsteel ten opzichte van de bladschijf sterker ontwikkeld dan in de rest van het jaar. In het voorjaar zijn de bladeren dan weer karakteristiek ingesneden. Niet alle soorten vertonen de beschreven seizoensvariatie even sterk. We vinden het vooral bij de secties Vulgaria, Palustria en Spectabilia; minder bij de sectie Erythrosperma. Er bestaat ook variatie in blad in grote verschillende milieutypen. Op beschaduwde plaatsen zijn de bladeren weinig en niet karakteristiek ingesneden. Ook planten in sterk droge en beweide milieus kunnen afwijkende bladeren geven. Voor determinatie, waarbij bladkenmerken vaak belangrijk zijn, is het dan ook nodig om planten te verzamelen in de bloeitijd, tot half mei; daarna veranderen de bladeren snel van vorm en verliezen hun diagnostische waarde. Ook dient men geen planten van beschaduwde en sterk beweide plaatsen te verzamelen. Wil men toch niet te determineren planten van ongunsti- 62
11 Fig. 51. Variatie in bladvorm bij Taraxacum nordstedtii (sect. Specctabilia ); a: blad vanjuveniele plant; b : blad uit de bloeitijd (begin mei); c : blad uit de vruchttijd (beginjuni); d: blad uit de zomer (augustus). ge plaatsen op naam brengen, dan is het nodig om ze te kweken. Ten gevolge van de grote groeikracht geeft dit meestal weinig problemen. In de kas kan men voorjaarsfenotypen krijgen bij lage temperatuur (15 ) en lange dag. Samen met de seizoensvariatie in bladvorm gaat eenvariatie in aantal bladeren per plant. Het aantal bladeren is bij veel soorten in het voorjaar het na grootst, de bloei neemt het af, in de zomer is het het laagst en in het najaar neemt het weer toe en in het voorjaar is het weer maximaal. In sommige populaties in de duinen verliest 30% van de planten de bladeren geheel; ze leven dan ondergronds (T. lacistophyllum, Ietswaart & Vroman, 1974). Ook bij Vulgaria-soorten in graslanden komt dit verschijnsel voor. Vermoedelijk is deze seizoensvariatie, met consequenties voor ademhaling, transpiratie en fotosynthese, in de duinen een aanpassing tegen droogte en in graslanden tegen concurrentie. Niet alle kenmerken zijn even variabel. Zo zijn de kenmerken de van vrucht, de stijl en de stempel en het omwindsel relatief weinig variabel. 6. De bloeitijd In Hagendijk, Van Soest & Zevenbergen (1975) zijn gegevens te vinden over de bloeitijd van de soorten van de secties Obliqua, Erythrosperma, Palustria en Spectabi- 63
12 Fig. 52. De bloei van Taraxacum-populaties (sect. Vulgaria) in enkele biotopen. 64

13 lia. Volgens opgave bloeien de meeste soorten van de laatste week van april tot half mei. T. obliquum en T. agaurum bloeien 14 dagen later dan de meeste soorten, nl. de tweede helft van mei en T. tortilobum begint al in maart. Voor de sectie Vulgaria wordt bij de meeste soorten geen bloeitijd opgegeven. Een goede indruk nu van de bloei van Vulgaria-microspecies geeft fig. 52. Het betreft waarnemingen in vijf biotopen in de omgeving van Abcoude in 1980 en wel: in een steile op het zuiden georiënteerde bermhelling van een rijksweg, in een vlakke wegberm van een rijksweg, in een gazon, in een op het noorden georiënteerde steile bermhelling van een rijksweg en in een agrarisch weiland. Telkens is per week van dezelfde oppervlakte het aantal bloemhoofdjes geteld. De curven van fig. 52 geven het verloop van de bloei in dat jaar; het hoogste aantal getelde hoofdjes in het voorjaar is op 100% gesteld. Op iedere oppervlakte komen enkele microspecies die voor, per oppervlak verschillen. Met stippen zijn de maaidata aangegeven en de veebezetting met balkjes. Duidelijk blijkt dat het verloop van de bloei afhankelijk is van het milieu; de zuidhelling bloeit het eerst, al op 25 maart en het laaggelegen weiland het laatst. Er is een duidelijke hoofdbloeitijd in april-mei en in 3 biotopen is er een tweede bloeiperiode in septemberoktober. In de tussentijd staat de bloei evenwel niet stil en worden er nog steeds, zij het met lage frequentie, bloeiende planten gevonden. In de zachte winter van ging de bloei op de zuidhelling en het gazon ook in december en januari spaarzaam door. Opvallend is de continuebloei op het gazon met intensieve nabloei; een ditondanks ofliever dank zij de hoge maaifrequentie. De hoofdjes in de maanden geven november en december over het algemeen geen kiemkrachtig zaad meer. Op zonnige dagen in het voorjaar begint de bloei met het opengaan van de hoofdjes omstreeks 9.00 u. s morgens. Het maximale aantal geopende hoofdjes wordt omstreeks 14.00u. gevonden en daarna neemt het aantal af; omstreeks 20.00uur zijn alle hoofdjes weer gesloten. Op zonnige, warme zomerdagen is het bloeigedrag wat anders dan in het voorjaar. De hoofdjes gaan dan omstreeks 8.00 uur open, omstreeks uur zijn de meeste hoofdjes open en omstreeks uur zijn de hoofdjes weer gesloten. Het gehele gebeuren is sterk afhankelijk van licht, temperatuur en vocht. Op sombere regenachtige dagen gaan de hoofdjes niet open en beneden de 3 C blijven, ook bij zonnig weer, de hoofdjes meestal ook gesloten. 7. Over de herkomst en de oecologie van de Nederlandse Taraxacum-flora Wanneer we de literatuurgegevens over de herkomst van de Nederlandse secties en soorten van Taraxacum samenvatten, dan ontstaat het volgende fragmentaire en in sommige opzichten speculatieve beeld. Sectie Vulgaria Deze sectie is met beschreven soorten de grootste en vormenrijkste binnen Taraxacum. De Vulgaria komen van nature vnl. in de gematigde zone van Eurazië voor; door de mens zijn ze evenwel over alle continenten verspreid (Dole, 1977; Van Soest, 1958). Het grootste deel der soorten is triploïd en agamosperm, behalve in Centraal- Europa, waar veel diploide taxa voorkomen. Voor Nederland worden tot nu toe 153 soorten opgegeven; dit is niet het totale aantal 65
 zouden de soorten van de sectie Vulgaria na de laatste ijstijd ontstaan zijn en wel uit Spectabilia-populaties, die de over ijsvrije vlakten vanuithet noorden naar het zuiden")
14 soorten binnen onze landsgrenzen; waarschijnlijk zijn er nog meer. Volgens Doll (1977) zouden de soorten van de sectie Vulgaria na de laatste ijstijd ontstaan zijn en wel uit Spectabilia-populaties, die de over ijsvrije vlakten vanuithet noorden naar het zuiden migreerden. Bij deze kolonisatie zouden door oecologische en geografische differentiatie de Vulgaria zijn ontstaan. Het is hierbij waarschijnlijk dat ook er hybridisaties hebben plaats gehad tussen Spectabilia-soorten en soorten van de noordelijke sectie Boreigena. Ook hybridisaties met reeds ontstane Vulgaria moeten niet uitgesloten worden geacht. De Vulgaria hebben zich postglaciaal snel over vrijwel geheel Europa verspreid met uitzondering van de meest noordelijke en meest zuidelijke delen.de Vulgaria behoren in deze zienswijze tot de jongste secties van Taraxacum. Richards (1972) heeft over de herkomst der Vulgaria een geheel andere theorie. Volgens deze auteur zouden de sexuele voorouders van de Vulgaria in Zuid-Europa de ijstijden in isolatie hebben overleefd. In deze periode werden geen isolatiemechanismen opgebouwd. Bij het stijgen van de temperatuurvolgde migratie, waarbij de soorten met elkaar in contact kwamen en hybridiseerden. Er zou zich daarna een zeer grote diploïde sexuele hybridenzwerm noordwaarts in Europa hebben verplaatst, die ten slottein contact kwam met triploïde agamosperme soorten van de oudere noordelijke secties Ceratophora en Arctica. Weer zou hybridisatie het gevolg zijn geweest en er ontstond zo een nieuwe grote hybridenzwerm, die. nu als het ware bevroren werd op agamosperm triploïd niveau. Uit de enorme hoeveelheid werden genotypen door natuurlijke selectie de slecht aangepaste geëlimineerd. Door deze verarming werden de miljoenen genotypen gereduceerd tot enkele duizenden. Ieder van deze genotypen plantte zich voort asexueel, agamosperm, en vormde zo klonen. Deze klonen, die genetisch van elkaar geïsoleerd waren, vertoonden morfologische differentiatieen hadden zich oecologisch gespecialiseerd. Door taxonomen worden ze nu als microspecies onderscheiden. De gedachte van door agamospermie bevroren hybridenzwermen, waaruit zich na natuurlijke selectie en adaptatie de microspecies van agamosperme groepen zouden hebben ontwikkeld, is niet nieuw. Gustafsson bespreekt deze veronderstelling in 1947 en wijst haar als gedeeltelijk onjuist van de hand. Hij geeft uitvoerig aan datook binnen agamosperme groepen het ontstaan van nieuwe genetische variatie mogelijk is, o.m. door hybridisatie en polyploïdisatie partieel sexuele van vormen en door mutatie en autosegregatie (zie Gustafsson, 1946). Hybriden tussen verschillende secties van Taraxacum zijn in Centraal-Europa in de natuur gevonden zij zijn ook in de proeftuin en gemaakt, zodat hybridisatie als bron van nieuwe variatie niet alleen meer theorie is (vgl. Den Nijs & Sterk, 1980). Sedert de ontdekking van het op uitgebreide schaal voorkomen van diploïdie en sexualiteit bij de Vulgaria in Midden-Europa, is het statische beeld van de bevroren hybridenzwerm voor een belangrijk deel wel van de baan. Zij is evenwel niet totaal van de baan, omdat het heel goed denkbaar is, dat in bepaalde delen van het areaal perioden van agamospermie afwisselen met perioden van gehele of gedeeltelijke sexualiteit en hybridisatie ofdat agamospermegeneraties afwisselen met sexuele, zodat er incidenteel hybridenzwermen optreden. In Noordwest-Europa overheerst de agamospermie, maar wellicht zullen hier bij nader onderzoek wel de bovenaangeduide en door Gustafsson beschreven processen ook worden aangetoond, zij het op zeer bescheiden schaal. Zoals reeds eerder opgemerkt, 66
.")
15 hebben Sorensen& Gudjonsson (1946) bewezen, dat er terugkeer naar sexualiteit mogelijk is bij triploïde agamosperme soorten, wanneer er chromosoomverlies optreedt. De microspecies van de sectie Vulgaria vinden we vnl. in storingsmilieus, de op meest uiteenlopende vaak anthropogene habitats (Londo, 1978; Oosterveld, 1978). Ze komen veel in voor vegetaties, die behoren tot het Lolio-Plantaginion en het Agropyro- Rumicion crispi en verder vinden we ze ook in het Arrhenatherionelatioris (Westhoff & Den Held, 1969). Sommige microspecies hebben voorkeur voor weinig bemeste of zilte of droge of vochtige biotopen (vgl. Oosterveld, 1978). Vulgaria-soorten kunnen zeer massaal optreden in sterk bemeste en beweide graslanden. Over de sectie Vulgaria bestaat danook zeer uitgebreide agrarisch georiënteerde literatuur.hierin worden geen microspecies onderscheiden, maar er is meestal sprake de van grote soort,,t. officinale. Vermoedelijk zijn bij deze soort ook meestal de Palustria- en Spectabiliasoorten gerekend (Kruijne, De Vries & Mooi, 1967). In de cultuurgraslanden is het aantal plantesoorten per perceel sedert het op uitgebreide schaal gebruiken van kunstmest sterk afgenomen, van gemiddeld soorten tot 1015 soorten; opvallend is nu dat het aantalindividuen (en microspecies?) van de Vulgaria daarentegen sterk is toegenomen. Per perceel kunnen wel tot 20 microspecies van Taraxacum voorkomen. Door het agrarisch beheer, hoge bemesting, sterke beweiding, ontwatering en inzaaien met bepaalde grasmengsels zijn veel graslanden tot onnatuurlijke biotopen geworden. In deze kunstmatige weilanden, die een grote oppervlakte binnen Nederlandenin West-Europa innemen, heeft zich een grote vormenrijkdom Vulgaria-microspecies ontwikkeld. van Het zal o.m. gaan om microspecies, die vroeger zelden voorkwamen, b.v. op en rondom boerenervenen die nu een grote verspreiding hebben gekregen, maar vermoedelijk ook om microspecies, die recent ontstaan en geëvolueerd zijn. Veel van deze microspecies zullen het karakter van agro-ecotypen hebben, d.w.z. het zijn oecologische varianten, die alleen in de agrarische sfeer voorkomen en die aangepast zijn aan agrarisch beheer. Buiten de landbouwsfeer worden niet ze gevonden. Deze agro-ecotypen (microspecies) verdwijnen bij verschraling volgens een bepaald patroon, z.g. verdwijnreeksen (Oosterveld, 1978). Vóór de agrarische revolutie zullen Vulgaria-soorten veel minder talrijk geweest zijn en in de weilanden zullen, zeker ook als gevolg van minder goede waterbeheersing, vooral de Vulgaria-soorten met voorkeur voor vochtige voedselarme biotopen en Palustria- en Spectabilia-soorten aanwezig zijn geweest. Sectie Palustria Deze sectie omvat + 50 soorten en is verspreid in Europa. Het is evenals de Vulgaria een typische laaglandsectie, die waarschijnlijk ontstaan is in Zuidoost-Europa vóór of tijdens de laatste ijstijd (Dole, 1977). Het merendeel der soorten is triploïd en tetraploïd; er zijn ook pentaploïde en aneuploïde taxa gevonden. Voor Nederland worden 10 soorten vermeld. Volgens Malecka (1965, 1973) zijn de Palustria hooggedifferentieerde taxa, bestaande uit allo- en autoploïden en veel taxa zouden facultatief agamosperm zijn. Het zijn vooral vochtminnende soorten, die sterke beweiding en bemesting niet kunnen verdragen. Sommige soorten hebben voorkeur voor zilte standplaatsen (zie Oosterveld, 1978). In Nederland komen ze vooral voor in onbemeste hooilanden, 67
. De Palustria zijn in ons land zeldzame soorten geworden, die meestal alleen nog in natuurreservaten voorkomen.")
16 vaak op kleiige bodems in vegetaties van hetcaricion davallianae en het Calthion palustris (Westhoff & Den Held, 1969); verder in het Cirsio-Molinietum en in zilte graslanden in het Junceto-Caricetum extensae (Hagendijk, Van Soest & Zevenbergen, 1975). De Palustria zijn in ons land zeldzame soorten geworden, die meestal alleen nog in natuurreservaten voorkomen. Sectie Spectabilia Deze grote vormenrijke boreo-atlantische sectie omvat soorten wordt vnl. en in Noordwest-Europa gevonden; ze zou ontstaan zijn vóór de laatste ijstijd. In de sectie komen triploïden voor, maar ook veel tetraploïden en door dit laatste volgen ze de trend der arctische soorten, die veelal tetraploïd zijn (Doll, 1977). Ook een hexaploïd is gevonden. Voor Nederland worden 9 soorten vermeld, die weinig karakteristiek voor de Spectabilia zijn. Over de begrenzing van Spectabilia en Vulgaria bestaat geen eenstemmigheid. Richards (1972) rekent T. adamii en T. litorale tot de Spectabilia, terwijl Hagendijk, Van Soest & Zevenbergen (1975) deze soorten tot de Vulgaria rekenen. Het zijn in Nederland vochtminnende soorten van weinig ofniet bemeste graslanden, vaak op zure, venige bodems; sommige soorten hebben voorkeur voor brakke milieus en andere komen voor in vegetaties van het Molinioncoeruleae. Evenals de Palustria zijn de Spectabilia zeldzame soorten in Nederland geworden, die vrijwel alleen nog in natuurreservaten voorkomen. Sectie Erythrosperma Deze tamelijk grote sectie omvat ± 120 soorten en is vnl. in Europa verspreid; door de mens is ze ook buiten Europa gebracht. De sectie zou in het Alpengebied zijn ontstaan en behoortevenals de Vulgaria tot de jongere secties van Taraxacum; ze zou vnl. postglaciaal van oorsprong zijn (Doll, 1977; Van Soest, 1958). Veel soorten zijn triploïd, maar ook diploïde en tetraploïde komen voor. Vele taxa zijn naar schatting zeer jong, nietouder dan jaar. Hiertoe behoren T. proximum, T. disseminatum, T. scanicum en T. lacistophyllum. Voor Nederland worden 23 soorten genoemd. Een groot deel van de Nederlandse Erythrosperma behoort tot de zeer jonge taxa. Van de Erythrosperma vindenwe veel soorten op droge, natuurlijke en half-natuurlijke standplaatsen in de duinen, de op zandgronden in oostelijk Nederland en op kalkrijke gronden in Zuid-Limburg. Vele soorten komen voor in het Galio-Koelerion (Westhoff & Den Held, 1969). Sectie Obliqua Deze kleine sectie omvat totaal slechts 3 soorten met een verspreiding langs de kusten van de Noordzee en de Oostzee. Voor Nederland wordt één soort genoemd. Waarschijnlijk bestaan er nauwe relaties met de sectie Dissecta (T. dissectum). Vermoedelijk is T. obliquum afgeleid van T. dissectum uit de Pyreneeën in het postglaciaal. Zij zou in het subboreaal het huidige areaal in West-Europa bereikt hebben (Doll, 1977). T. obliquum is triploïd. De soort is gekenmerkt door het voorkomen in droge open, natuurlijke standplaatsen in duinen in het Taraxaco-Galietum maritimi (Westhoff & Den Held, 1969). 68

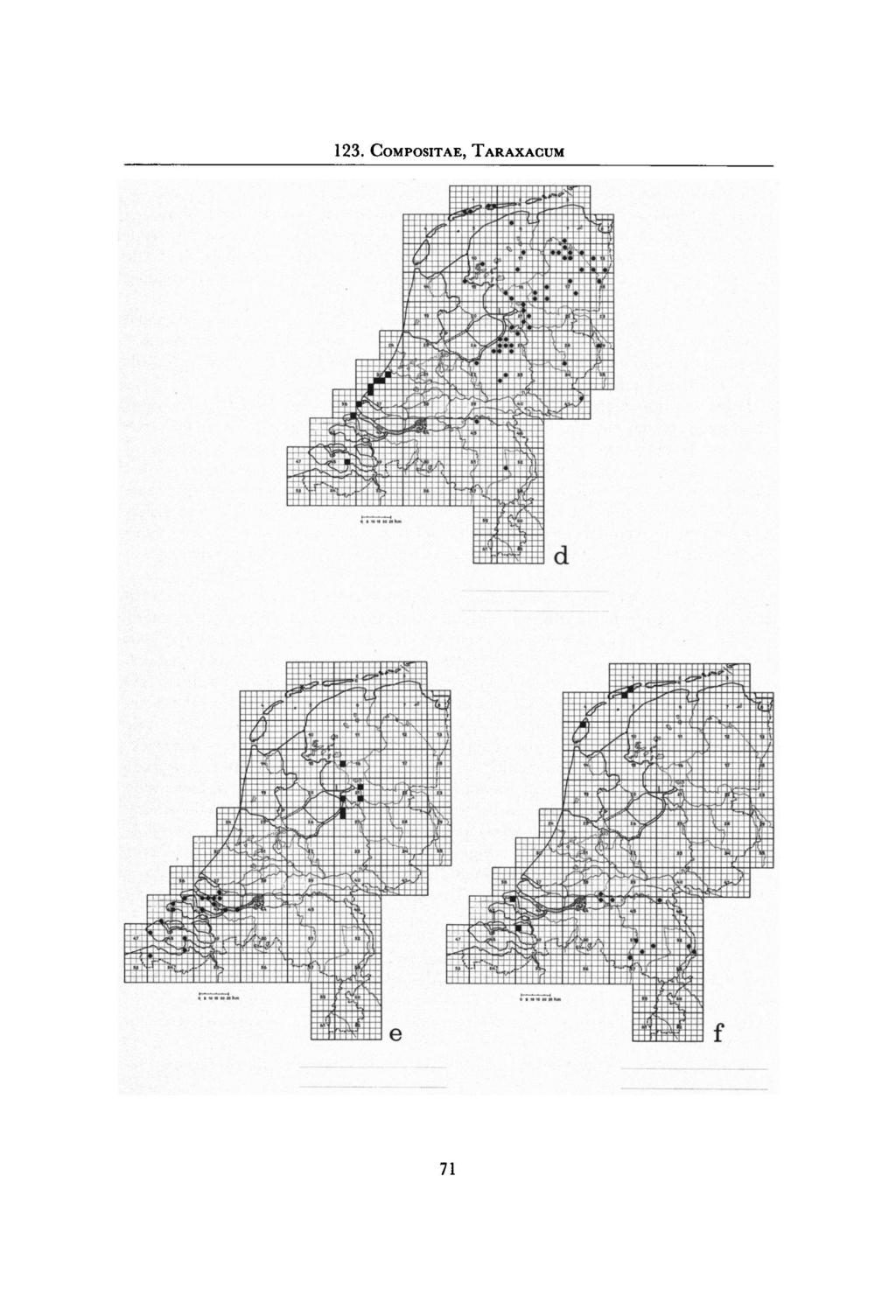
17 - alsook Samenvattend blijkt dat de Taraxacum-secties naar plaats en tijd naar voorouders (Doll,1977) - een zeer verschillende herkomst hebben. Ook oecologisch zijn de secties minof meer duidelijk verschillend. Van de autoecologie van de meeste soorten is niets bekend. 8. Plantengeografische aspecten Bij de bespreking van enkele plantengeografische aspecten van de Nederlandse Taraxaca zullen we ons in hoofdzaak beperken tot de sectie Vulgaria. Een gedetailleerde plantengeografische analyse in Westeuropees verband is nog niet mogelijk, omdat er over veel Nederlandse taxa nog geen gedetailleerde verspreidingsgegevens bekend zijn; bovendien is de Taraxacum-flora van de ons omringende landen nog onvoldoende onderzocht, met name die van West-Duitsland, België en Frankrijk. Wanneer we de verspreidingsgegevens van de Vulgaria-soorten uit de hiernavolgende Taraxacum-bewerking overzien, dan blijkt dat veel soorten een verspreiding hebben in het gebied dat Zuid-Scandinavië, Denemarken, Noordwest-Duitsland, Nederland, België, Zuid- Engeland en Noord-Frankrijk omvat. Weinige soorten gaan noordelijker en een klein aantal gaat zuidelijker en wel tot in Zwitserland. Een voorbeeld hiervan toont het areaal van T. hamatiforme (Fig. 53). Dit gebied is vnl. atlantisch vankarakter. Er is een zeer grote variatie in areaalgrootte. Sommige soorten omvatten het gehele boven genoemde gebied, andere een kleiner deel en sommige soorten zijn slechts van één ofenkele vindplaatsen bekend. De meeste soorten zullen in het postglaciaal Nederland zijn binnengekomen, bij vele soorten zal dit met natuurlijke verspreidingsmiddelen zijn gebeurd. Immigratie door menselijk transport zal evenwel ook zeer belangrijk zijn geweest en dat zal het zeker nog zijn, gezien het intensieve verkeer met het buitenland; dit geldt speciaal voor de soorten uit de vnl. anthropogene habitats. Fig. 53. De verspreidingvan Taraxacum hamatiforme (sect. Vulgaria) naar Van Soest, Hagendijk & Zevenbergen (1968). 69
, T.westhoffii(.);ƒ: T. brabanticum ( ), T. wijtmaniae( ). behoort tot de sect.")
18 Fig. 54. De verspreiding van enkele (Rijswijk); a: T. sellandii; b: T. raunkiaeri; c: T. multicorans ( ), T. texelense ( );d: T. atactum ( ), T. ordinatum ( );e: T. beeftinkii ( ), T. flevoense Taraxacum- soorten in Nederland naar gegevens van H. A. Zevenbergen ( ), T.westhoffii(.);ƒ: T. brabanticum ( ), T. wijtmaniae( ). behoort tot de sect. Erythrosperma, alle overige behoren tot de sect. Vulgaria. T. westhoffii 70
19 71
20 Sommige soorten komen in het gehele land algemeen voor, zoals T. sellandii, die een ruime oecologische tolerantieheeftmeteen voorkeur voor wegbermen en begraasde weilanden (fig. 54a). Ook buiten Nederlandis de soort ruim verspreid en wel van Zuid- Scandinavië tot Midden-Frankrijk. Het betreft hier een relatiefoude soort, die mede door zijn grote oecologische tolerantiein anthropogene habitats zijn ruime verbreiding heeft gekregen. Een andere ruim verspreide soort is T. raunkiaeri (fig. 54/;). Deze soort heeft een beperktere oecologische tolerantie dan T. sellandii; we vinden deze soort bij voorkeur op zandgrond. BuitenNederland komt T. raunkiaeri voor van Denemarken en Noordwest-Duitsland tot in Midden-Frankrijk en in Zuid-Engeland. Tegenover deze algemene soorten met een groot areaal staan de endemische meer kieskeurige soorten met een klein areaal zoals T. texelense (fig. 54c) en T. flevoense (fig. 54c). Beide soorten hebben vrij specifieke levenseisen; T. texelense is beperkt tot min of meer zilte gronden en T. flevoense tot vochtige graslanden behorende tot het Calthion en tot hooiweiden van het Fritillario-Alopecuretum pratensis. Het is waarschijnlijk, dat hun areaal zo klein is, omdat ze recent zijn onstaan. Een andere mogelijkheid is dat ze nog verder in en buitennederland voorkomen, maar doorhun zeldzaamheid nog niet zijn gevonden. Het is niet waarschijnlijk dat hun areaal een relict is van een vroeger veel groter areaal. Sommige jonge soorten breiden hun areaalin Nederland nog steeds uit; een voorbeeld hiervan is T. agaurum (Erythrosperma), die voorkomt in het Duindistrict van Noordwijk tot Voorne en die zich naar het noorden en zuiden uitbreidt (Hagendijk, Van Soest & Zevenbergen, 1975). Een extreem voorbeeld van een zeer klein endemisch areaal is T. westhoffii (Erythrosperma), die slechts op één plaats op Terschelling is gevonden en wel in een vochtig karrespoor in een duinvallei ten zuiden van de Koegelwieck (fig. 54c). Ook systematisch staat deze soort geïsoleerd. De bladvorm en de omwindselblaadjes lijken op die van Palustria-soorten, ook de oecologie wijst in de richting van de Palustria, de knobbeltjes op de buitenste omwindselblaadjes en de rode kleur van de vruchtjes zijn echter kenmerken van de Erythrosperma. Wellicht is T. westhoffii door hybridisatie van een Palustria- met een Erythrosperma-soort ontstaan. De hybride heeft zich niet verder kunnen uitbreiden en is vermoedelijk weer uitgestorven, in ieder geval is ze niet meer teruggevonden. T. westhoffii is niet de enige soort met kenmerken, die tussen de Palustria en Erythrosperma instaan. In Europa zijn er vier bekend; ze zijn verenigd in de z.g. Taraxacum langeanum-groep (Nordenstam & Van Soest, 1974). Sommige soorten lijken zeldzaam, omdat ze pas zeer recent beschreven zijn, zoals T. wijtmaniae, die in 1978 is gepubliceerd (fig. 54ƒ); waarschijnlijk is deze soort echter niet zeldzaamin het Deltagebied en het Waddengebied. Van de 153 Nederlandse Vulgaria-soorten worden er 25 alleen voor ons land opgegeven; 15 soorten hiervan zijn zeldzaam; 7 zijn vrij algemeen, 1 soort is algemeen en van 2 zijn er geen opgaven van zeldzaamheid. Sommige soorten hebben karakteristieke arealen in Nederland; enkele hiervan worden genoemd. T. atactum (fig. 54 d) komt vnl. in Noord-Nederland voor; deze soort treffen we verder aan in Noordwest-Duitsland en Zuid-Denemarken (Sahlin & Van Soest, 1970). 72
 vindenwe in Zuidwest-Nederland, daarbuiten waarschijnlijk niet. De soort is algemeen op zeeklei, zeldzamer op rivierklei.")
21 T. ordinatum (fig. 54öf) vinden we in het Duindistrict ten zuiden van Wassenaar tot in Zeeland, bij voorkeur aan de binnenduinrand op het zand; buiten Nederland is de soort in Engeland gevonden. T. multicolorans (fig. 54c) vindenwe in Zuidwest-Nederland, daarbuiten waarschijnlijk niet. De soort is algemeen op zeeklei, zeldzamer op rivierklei. We treffen haar aan op dijken en in graslanden (Hagendijk, Van Soest & Zevenbergen, 1972). T. beeftinkii (fig. 54e) is vrij algemeen in het Deltagebied, daarbuiten is ze nog niet gevonden. De soort komt voor op laag gelegen, iets ziltige graslanden (Hagendijk, Van Soest & Zevenbergen, 1972). T. brabanticum (fig. 54/) is een zeldzame soort van vochtige graslanden behorende tot het Calthion. We vindent. brabanticum in oostelijk Noord-Brabant en in Middenen Noord-Limburg, buiten Nederland nog bij Aken. 9. Zijn paardebloemesoorten wel goede soorten? Wanneer we over paardebloemesoorten spreken, dan bedoelen we in dit verband de soorten, die in Noord-Europa en ook in ons land voorkomen en die zich asexueel, agamosperm, voortplanten. Het is niet de bedoeling het algemene probleem van de classificatie en systematiek van agamosperme taxa te behandelen. Over deze vraagstukken bestaat een uitvoerige literatuur, waarnaarwordt verwezen (Doll, 1972). We willen volstaan met enkele opmerkingen. Bij Taraxacum-soorten die zich agamosperm voortplanten, is de nakomelingschap genetisch identiekaan de moederplant (behoudens mutaties). Er ontstaan op deze wijze agamosperme afstammingsreeksen, die in feite klonen zijn. Zeer waarschijnlijk bestaan de soorten van Taraxacum uit één kloon of een mengsel van klonen. In de praktijk nu blijkt dat dezeklonenof mengsels van klonen morfologisch vaak vanelkaar te onderscheiden zijn en dat zij een eigen oecologische preferentie en tolerantiehebben (althans daarvoor zijn veel aanwijzingen: van Loenhoud & Duyts, 1981). Bovendien is van doorslaggevende betekenis, dat de soorten genetisch van elkaar geïsoleerd zijn. Deze eigenschappen vormen de biologische achtergrond voor het onderscheiden van taxonomische soorten. Omdat deze soorten meestal een smal variatietraject hebben, worden ze vaak microspecies (kleine soorten) genoemd. Het is een algemene karakteristiek van agamosperme taxa (microspecies) in het plantenrijk dat ze talrijk zijn en veel op elkaar lijken; dit is dus niet een speciaal kenmerk van de taxa van Taraxacum. Voor het verkrijgen van een goed inzicht in de microspecies van Taraxacum, nog een enkele opmerking over de biologische soort. Wanneer er genetische isolatiemechanismen tussen populaties bestaan, danbehoren deze populaties tot verschillende biospecies. Dit is evolutionair van groot belang, omdat deze populaties door hun genetische isolatie in de evolutie dan hun eigen weg gaan. Biologische soorten vinden we bij z.g. biparentale voortplanting, waarbij de voortplanting plaats vindt via twee ouders. Door meiose en bevruchting (via kruisbestuiving) komt dan de genenuitwisseling tot stand. Bij uniparentale voortplanting, waarbij één ouder nakomelingen voortbrengt, zoals bij obligate zelfbestuiving en obligate agamospermie, is er géén genenuitwisseling en de nakomelingschappen van individuele ouders zijn genetisch van elkaar geïsoleerd. 73
22 Obligaat agamosperme taxa, zoals de Taraxaca in Noord-Europa meestal zijn, vormen dan ook door het ontbreken van genenuitwisseling geen biologische soorten. Biologisch zijn biparentale soorten en uniparentale soorten geheel verschillende systemen; beide vormen ze echterwel taxonomische soorten. Grant (1971) heeft uniparentale en biparentale soorten verenigd in z.g. evolutionaire soorten. Wanneer we ons afvragen of de Taraxacum-species wel goede taxonomische soorten zijn en geschikt voor algemeen bruikbaar wetenschappelijk onderzoek, dan hangt het antwoord vnl. afvan de practische herkenbaarheid der soorten, aangenomen dat de eerder genoemde biologische achtergrond der soorten gezond is. Met name is de vraag belangrijk of de soorten morfologisch voldoende vanelkaarafte grenzen zijn, waardoor identificatiemet een determinatietabel, eventueelondersteund met een vergelijkingscollectie van herbariumplanten, mogelijk is. Tot nu toe zijn er twee typen van Taraxacumsoorten in de literatuurbekend. De oudste zijn de grote z.g. Linneaanse soorten, o.m. onderscheiden door Handel-Mazzetti (1907). Deze soorten omvatten meestal een aantal microsoorten, soms zelfs wel een gehele sectie, zoals T. vulgare Schrank. Ze zijn echter zeer heterogeen, hebben soms 2 3 chromosoomgetallen en omvatten oecologisch en morfologisch zeer verschillende groepen. Deze soorten worden door de meeste auteurs niet als goede soorten beschouwd. Naast deze grote soorten zijn er de al vaak genoemde kleine of microsoorten, die nu door veel taxonomen worden gebruikt. Deze soorten hebbenals nadeeldat ze zoveel op elkaar lijken, dat meestalalleen specialisten met zekerheid de planten kunnen identificeren. Dit geldt althans voor veel van deze soorten, met name van de sectie Vulgaria. In de sectie Erythrosperma zijn de taxonomische moeilijkheden minder groot dan in de andere Nederlandse secties. Het antwoord op de vraag of de microsoorten van Taraxacum wel goede taxonomische soorten zijn, zal genuanceerd moeten worden beantwoord. Van belang hierbij is, dat er gradaties bestaan in herkenbaarheidder microsoorten. Sommige soorten zijn goed herkenbaar, b.v. T. obliquum en T. adamii; de meeste echter vertonen onderling zeer grote overeenkomsten en zijn eerst herkenbaar nadat men zich goed in de heeft groep ingewerkt. In het algemeen zullen we moeten zeggen, dat de Taraxacum-soorten voor een algemeen bruikbareclassificatie niet geschikt zijn. Richards (1972) schat dat met zijn,,taraxacumflora ofthe British Isles botanici zonder speciale Taraxacum-kennis 80% van de planten op naam kunnen brengen. Voor Nederland zal dit optimistische percentage zeker niet hoger liggen. Essentieel bij determinaties van paardebloemen is een goed referentieherbarium. We zullen verder bij identificatie nog met een ander probleem rekening moeten houden en wel dat een bepaald percentage der planten uit de natuur niet op naam te brengen is, ook niet door specialisten. Hagendijk, Van Soest & Zevenbergen noemen hiervoor in de hierna volgende inleiding tot hun bewerking van de Vulgaria een aantal oorzaken, waarnaar wordt verwezen. Als mogelijke oorzaak van classificatiemoeilijkheden moet, gezien de recente ontdekking van sexuele planten, zeker ook hybridisatie worden genoemd. De enige manier om niet te determineren paardebloemen uit bij voorbeeld sterk begraasde weilandenof van beschaduwde plaatsen toch op naam te brengen is ze te kweken en dit is dan ook een werkwijze, die door veel taraxacologen is gevolgd. Voor specialisten zijn vele microspecies, dieworden genoemd in de Flora Neerlandica, gezien 74
23 & de vele morfologische studies die zijn verricht en het uitgebreide kweekwerk dat is gedaan, goede soorten, in die zin dat ze morfologisch herkenbaar zijn en dat ze constant zijn in de tijd. Voorzichtigheid blijft evenwel geboden met betrekking tot het laatste punt. Onvoldoende is nog onderzocht in hoeverre de geconstateerde sexualiteit in de agamosperme populaties voorkomt. Ook deze wanneer met lage frequentie aanwezig is, dankan ditten gevolge van hybridisatie toch grote gevolgen hebbenvoor de identiteit der microspecies en het ontstaan van nieuwe varianten en taxa. Hierbij komt dat van veel Vulgaria-soorten het aantal exemplaren, datin de Nederlandse herbaria aanwezig is, nog betrekkelijk klein is, waardoor de variatie per microsoort voor ons land nog onvoldoende bekend is. Nader onderzoek aan meer exemplaren zou kunnen uitwijzen dat de verschillen tussen sommige soorten gaan vervagen, waardoor ze in één taxon geclassificeerd moeten worden. Het is ook niet onwaarschijnlijk, datin een aantal gevallen standplaatsmodificaties als aparte soorten beschreven zijn. Nader onderzoek blijft dan ook noodzakelijk. Voor Taraxacum geldt nog de bijzondere moeilijkheid, dat microspecies moeilijk in goed te definiëren groepen samen te vatten zijn; dit is een gevolg van hun reticulate (netvormige) verwantschappen. Het is dan ook nog vrijwel niet mogelijk om tussen de microspecies en de sectie een taxonomisch niveau in te voegen, of om microspecies te classificeren als subspecies van meer omvattende species. Tenslotte een opmerking over de infraspecifieke variatie. Uit de literatuur (o.m. Doll, 1972; Hagendijk, Van Soest & Zevenbergen, 1975) blijkt dat ook deze bij Taraxacum voorkomt. Het is van belang hierbij te bedenken dat niet iedere levensvatbare mutatie, die bij een microsoort voorkomt, de oorzaak is van het ontstaan van een nieuwemicrosoort. Evenals bij sexuele soorten vinden we ook bij agamosperme soorten een infraspecifieke variatie, die vnl. oecologisch en geografisch is bepaald. Literatuur Bosch, W. B., Chromosoomtellingen en vruchtmorfologie bij Taraxacum. Intern rapp. Hugo de Vries-lab. 78, Amsterdam. Candolle, A. P. de, Prodromus systematis naturalis Regni Vegetabilis 7, p Dole, R., Die apomiktische Art und ihre Beziehungzur Evolution. In W. Vent, Widerspiegelung der Binnenstruktur und Dynamik der Art in der Botanik, Symp. Arb.gem. Phytotax. Biol. Ges. D.D.R., p , Revision der sect. Erythrosperma Dahlst. emend. Eind. f. der Gattung Taraxacum Zinn. Fedd. Rep. 84, p ,, Die Gattung Taraxacum. Lutherstadt Grundrisz der Evolution der Taraxacum Zinn. Gattung Thesis. Berlin. Faegri, K. & L. van der Pijl, Principles of pollination ecology. Oxford. Fengler, A. G., Een biosystematische studie aan Taraxacum-soorten in graslanden op Intern Terschelling. rapp. Hugo de Vries-lab. 61, Amsterdam. A. A. Sterk, Taraxacum wijtmaniae, spec. nov. Gorteria 9, p Grant, V., Plant speciation. London. Gustafsson, A., Apomixis in higher plants, I. The mechanism of apomixis. Act. Univ. Lund. N.S. 42, p
. Flora Neerl.")
24 & &, Apomixis in higher plants. II. The causal aspects of apomixis. Act. Univ. Lund. N.S. 43, p Hagendijk, A., P. Oosterveld & H. A. Zevenbergen, Taraxacum vansoestii, spec. nov. Gorteria 9, p , J. L. van Soest & H. A. Zevenbergen, Neue Taraxacumarten der Niederlande. Act. Bot. Neerl. 21, p ,, Taraxacum (behalve sectie Vulgaria). Flora Neerl. IV (9), p Handel-Mazzetti, H. von, Monographie der Gattung Taraxacum. VVien. Heukels-Wachter, Geïllustreerde Schoolflora voor Nederland, ed. 11. Groningen. Hou-Liu, S. Y., The chromosome counts of some Taraxacum species. Act. Bot. Neerl. 12, p Ietswaart, J. H. & M. Vroman, Seizoensveranderingen bij bladeren van Taraxacum. Gorteria 7, p Kruijne, A. A., D. M. devries & H. Mooi, Bijdrage tot de oecologie van de Nederlandse graslandplanten. Versl. landbouwk. onderz Wageningen. Linnaeus, C., Species Plantarum. Stockholm. Loenhoud, P, J. van & H. Duyts, A comparative study of the germination ecology of some microspecies oftaraxacum Wigg. Act. Bot. Neerl. 30, p Londo, G., Over het gedrag in ruimte en tijd van Taraxacum en Plantage. Gorteria 9, p Malecka,J., Embryologicalstudies in Taraxacum palustre. Act. Biol. Crac. 8, p , Problems of the mode of reproduction in microspecies oftaraxacum section Palustria Dt. Act, Biol. Crac. 16, p Mooren, J., Over de Taraxacum-flora van Nederlandse graslanden. Intern rapp. Hugo de Vries-lab., Amsterdam, in druk. Nordenstam, S. & J. L. van Soest, Taraxacum leporinum, spec. nov. Bot. Not. 127, p Nys, J. C. M. den & A. A. Sterk, Cytogeographical studies oftaraxacum sect. Taraxacum ( sect. Vulgaria) in Central Europe. Bot. Jahrb. Syst. 101, p , H. van der Hammen, Cytological and ecological notes the on Taraxacumsections Erythrosperma and Obliqua of the Coastal area of the Nctherlands. Act. Bot. Neerl. 27, p Oosterveld, P., De indicatiewaarde van het genus Taraxacum voor het beheer van graslanden. Gorteria 9, p Richards, A. J., The Taraxacum flora of the British Isles. Watsonia 9, Suppl., p , The origin oftaraxacum agamospecies. Bot. Journ. Linn, Soc. 66, p & P. D. Sell, Taraxacum Wigg., in T. G. Tutin c.s., Flora Europaea 4, p Cambridge. Rutishauser, A., Fortpflanzungsmodus und Meiose apomiktischer Blütenpflanzen. Protoplasmologia VI, F3. Sahlin, C. I. & J. L. van Soest, Taraxacum atactum, spec. nov. Act. Bot. Neerl. 19, p Soest, J. L. van, Het geslacht Taraxacum in Nederland I. Ned. Kruidk. Arch. 49, p , Het geslacht Taraxacum in Nederland II.Ned. Kruidk. Arch. 52, p ,, Taraxacum Sectio Vulgaria Dt. in Nederland I. Act, Bot. Neerl. 4, p The phytogeography oftaraxacum with special reference to Europe. Blumea Suppl. 4, p , Taraxacum Sect. Palustria Dt. Act. Bot. Neerl. 14, p , A. Hagendijk & H. Zevenbergen, Taraxacum kernianum, nov. spec. Gorteria 4, p
indeling bleef gezaghebbend tot de laatste eeuwwisseling. Het was ook het uitgangspunt
 Classificatie en sexualiteit bij Taraxacum door A.A. Sterk en J.C.M. den Nijs (Vakgroep Bijzondere Plantkunde, Amsterdam) I. Inleiding Lange tijd is gedacht dat het geslacht Taraxacum soortenarm en eenvormig
Classificatie en sexualiteit bij Taraxacum door A.A. Sterk en J.C.M. den Nijs (Vakgroep Bijzondere Plantkunde, Amsterdam) I. Inleiding Lange tijd is gedacht dat het geslacht Taraxacum soortenarm en eenvormig
kijken; de tocht bracht een Op 10 mei 1966maakte ik met de heren P. A. Bakker, C. Boot enchr. van Leeuwen Taraxacum euryphyllum door
 Taraxacum euryphyllum (Dahlst.) Christ. in Nederland door J.L. van Soest ('s-gravenhage) Op 10 mei 1966maakte ik met de heren P. A. Bakker, C. Boot enchr. van Leeuwen een excursie naar de reservaten op
Taraxacum euryphyllum (Dahlst.) Christ. in Nederland door J.L. van Soest ('s-gravenhage) Op 10 mei 1966maakte ik met de heren P. A. Bakker, C. Boot enchr. van Leeuwen een excursie naar de reservaten op
Evolutie van de generatiewisseling
 Evolutie van de generatiewisseling Oefen- en zelftoetsmodule bij hoofdstukken 29 en 30 van Campbell november 2006 Introductie Met behulp van dit zelfstudie-onderdeel oefen je de termen en begrippen die
Evolutie van de generatiewisseling Oefen- en zelftoetsmodule bij hoofdstukken 29 en 30 van Campbell november 2006 Introductie Met behulp van dit zelfstudie-onderdeel oefen je de termen en begrippen die
Aan de Minister van Volkshuisvesting, Ruimtelijke Ordening en Milieubeheer Mevrouw dr. J.M. Cramer Postbus GX Den Haag
 Aan de Minister van Volkshuisvesting, Ruimtelijke Ordening en Milieubeheer Mevrouw dr. J.M. Cramer Postbus 30945 2500 GX Den Haag DATUM 2 juli 2008 KENMERK CGM/080702-01 ONDERWERP Inschaling van handelingen
Aan de Minister van Volkshuisvesting, Ruimtelijke Ordening en Milieubeheer Mevrouw dr. J.M. Cramer Postbus 30945 2500 GX Den Haag DATUM 2 juli 2008 KENMERK CGM/080702-01 ONDERWERP Inschaling van handelingen
Enige opmerkingen. afwezigheid van uitlopers (stolonen), de aanwezigheid van één of van twee rijen haren bij
, de aanwezigheid van één of van twee rijen haren bij") Enige opmerkingen over de voortplantingswijze van Hieracium pilosella L. en enkele verwante taxa door Th.W. J. Gadella (Vakgroep Populatie- en Evolutiebiologie, Utrecht) Het omvangrijke Hieracium genus
Enige opmerkingen over de voortplantingswijze van Hieracium pilosella L. en enkele verwante taxa door Th.W. J. Gadella (Vakgroep Populatie- en Evolutiebiologie, Utrecht) Het omvangrijke Hieracium genus
Evolutie van het Melkviooltje en verwanten door hybridisatie en polyploïdie
 Evolutie van het Melkviooltje en verwanten door hybridisatie en polyploïdie De systematiek is de wetenschap binnen de biologie die zich bezighoudt met inventariseren, identificeren van de verwantschapsrelaties
Evolutie van het Melkviooltje en verwanten door hybridisatie en polyploïdie De systematiek is de wetenschap binnen de biologie die zich bezighoudt met inventariseren, identificeren van de verwantschapsrelaties
Dahlst. em. Lindb. f. door. toestand neutraal bruin (notenhout, donker. lc Vruchten geelachtig gebakken steenrood, terra-cotta oflicht zalmkleurig.
 Fig Korte determinatietabel voor de Nederlandse soorten van Taraxacum sect Erythrosperma Dahlst em Lindb f door JL van Soest ('s-gravenhage) la Vruchten strokleurig (tot bleekbruin) 2 Vgl ook T obliquum
Fig Korte determinatietabel voor de Nederlandse soorten van Taraxacum sect Erythrosperma Dahlst em Lindb f door JL van Soest ('s-gravenhage) la Vruchten strokleurig (tot bleekbruin) 2 Vgl ook T obliquum
Oplossingen Biologie van 2000
 www. Oplossingen Biologie van 2000 Disclaimer: Alle uitwerkingen zijn onder voorbehoud van eventuele fouten. Er is geen enkele aansprakelijkheid bij de auteur van deze documenten. Vraag 1 Om deze vraag
www. Oplossingen Biologie van 2000 Disclaimer: Alle uitwerkingen zijn onder voorbehoud van eventuele fouten. Er is geen enkele aansprakelijkheid bij de auteur van deze documenten. Vraag 1 Om deze vraag
3 Rundveefokkerij Melkproductiecontrole Selectie Fokwaardeschatting Inseminatieplannnen 69 3.
 Inhoud Voorwoord 5 Inleiding 6 1 Veiligheidsvoorschriften 9 1.1 Genen en hun vererving 9 1.2 Genotype en fenotype 14 1.3 Erfelijke gebreken 18 1.4 Genfrequenties 25 1.5 Afsluiting 27 2 Fokmethoden 28 2.1
Inhoud Voorwoord 5 Inleiding 6 1 Veiligheidsvoorschriften 9 1.1 Genen en hun vererving 9 1.2 Genotype en fenotype 14 1.3 Erfelijke gebreken 18 1.4 Genfrequenties 25 1.5 Afsluiting 27 2 Fokmethoden 28 2.1
Mitose is een ander woord voor gewone celdeling. Door gewone celdeling blijft het aantal chromosomen in lichaamscellen gelijk (46 chromosomen).
.") Samenvatting door M. 1493 woorden 28 februari 2014 5 5 keer beoordeeld Vak Methode Biologie Biologie voor jou Genotype en fenotype Veel eigenschappen zijne erfelijk. Je hebt deze eigenschappen geërfd van
Samenvatting door M. 1493 woorden 28 februari 2014 5 5 keer beoordeeld Vak Methode Biologie Biologie voor jou Genotype en fenotype Veel eigenschappen zijne erfelijk. Je hebt deze eigenschappen geërfd van
doorgaans met deperceelgrenzen overeen te komen enhet ligt dan ook voor de hand om te
 De indicatiewaarde van het genus Taraxacum voor het beheer van graslanden door P. Oosterveld (Rijksinstituut voor Natuurbeheer, Leersum) Inleiding Wie zich in april-mei doorhet Nederlandse landschap beweegt,
De indicatiewaarde van het genus Taraxacum voor het beheer van graslanden door P. Oosterveld (Rijksinstituut voor Natuurbeheer, Leersum) Inleiding Wie zich in april-mei doorhet Nederlandse landschap beweegt,
Alsemambrosia Ambrozijn Ambrosia artemisiifolia
 Alsemambrosia Ambrozijn Ambrosia artemisiifolia Een exoot PlantenWerkGroup Scousele Sonja Reps 11/10/2017 2 Beschrijving: Alsemambrosia is een éénjarige plant die pas sinds 1875 voor het eerst in ons land
Alsemambrosia Ambrozijn Ambrosia artemisiifolia Een exoot PlantenWerkGroup Scousele Sonja Reps 11/10/2017 2 Beschrijving: Alsemambrosia is een éénjarige plant die pas sinds 1875 voor het eerst in ons land
Bouw zaadplanten. CC Naamsvermelding 3.0 Nederland licentie.
 Auteur VO-content Laatst gewijzigd 16 December 2016 Licentie CC Naamsvermelding 3.0 Nederland licentie Webadres http://maken.wikiwijs.nl/87623 Dit lesmateriaal is gemaakt met Wikiwijs Maken van Kennisnet.
Auteur VO-content Laatst gewijzigd 16 December 2016 Licentie CC Naamsvermelding 3.0 Nederland licentie Webadres http://maken.wikiwijs.nl/87623 Dit lesmateriaal is gemaakt met Wikiwijs Maken van Kennisnet.
Celdeling vmbo-kgt12. CC Naamsvermelding 3.0 Nederland licentie.
 Auteur VO-content Laatst gewijzigd 14 July 2016 Licentie CC Naamsvermelding 3.0 Nederland licentie Webadres http://maken.wikiwijs.nl/62460 Dit lesmateriaal is gemaakt met Wikiwijs Maken van Kennisnet.
Auteur VO-content Laatst gewijzigd 14 July 2016 Licentie CC Naamsvermelding 3.0 Nederland licentie Webadres http://maken.wikiwijs.nl/62460 Dit lesmateriaal is gemaakt met Wikiwijs Maken van Kennisnet.
Inhoud. Voorwoord 5. Trefwoordenlijst 113. Inhoud
 Inhoud Voorwoord 5 6 Steeds meer planten! 9 6.1 Geslachtelijke variaties 9 6.2 Van eicel tot zaad 11 6.3 Allemaal gelijk 14 6.4 De juiste behandeling 24 6.5 Verschillende bollen en knollen 28 6.6 Afsluiting
Inhoud Voorwoord 5 6 Steeds meer planten! 9 6.1 Geslachtelijke variaties 9 6.2 Van eicel tot zaad 11 6.3 Allemaal gelijk 14 6.4 De juiste behandeling 24 6.5 Verschillende bollen en knollen 28 6.6 Afsluiting
Bloemen. Duinroosje. Bloemen
 Duinroosje Bloemtypen. Eén- en tweeslachtige bloemen. Tweeslachtige bloemen. De bloem heeft twee geslachten, d.w.z meeldraden en stampers zitten in één bloem bij elkaar. Eénslachtige bloemen. De bloem
Duinroosje Bloemtypen. Eén- en tweeslachtige bloemen. Tweeslachtige bloemen. De bloem heeft twee geslachten, d.w.z meeldraden en stampers zitten in één bloem bij elkaar. Eénslachtige bloemen. De bloem
memo gemeente Lingewaard, Dion Steenbergen , hoofdlijnen mitigatieplan kleine wolfsmelk Houtakker II, Bemmel
 memo aan: van: gemeente Lingewaard, Dion Steenbergen SAB, Eric Verkaik c.c.: datum: 15 september 2017 betreft: 160472, hoofdlijnen mitigatieplan kleine wolfsmelk Houtakker II, Bemmel INLEIDING Aan de Houtakker
memo aan: van: gemeente Lingewaard, Dion Steenbergen SAB, Eric Verkaik c.c.: datum: 15 september 2017 betreft: 160472, hoofdlijnen mitigatieplan kleine wolfsmelk Houtakker II, Bemmel INLEIDING Aan de Houtakker
Voorbeelden van indeling van levensstadia of levensfasen bij verschillende planten en plantengroepen.
 Voorbeelden van indeling van levensstadia of levensfasen bij verschillende planten en plantengroepen. Gatsuk et al. (1980) geven de volgende standaardindeling voor in Rusland onderzochte soorten: Ontogenetische
Voorbeelden van indeling van levensstadia of levensfasen bij verschillende planten en plantengroepen. Gatsuk et al. (1980) geven de volgende standaardindeling voor in Rusland onderzochte soorten: Ontogenetische
Jongetje of meisje? hv12. CC Naamsvermelding-GelijkDelen 3.0 Nederland licentie.
 Auteur VO-content Laatst gewijzigd 24 October 2016 Licentie CC Naamsvermelding-GelijkDelen 3.0 Nederland licentie Webadres http://maken.wikiwijs.nl/62534 Dit lesmateriaal is gemaakt met Wikiwijs Maken
Auteur VO-content Laatst gewijzigd 24 October 2016 Licentie CC Naamsvermelding-GelijkDelen 3.0 Nederland licentie Webadres http://maken.wikiwijs.nl/62534 Dit lesmateriaal is gemaakt met Wikiwijs Maken
Anatomie en Morfologie. Wortel. Plantencel. Stengel. Delen van een zaadplant
 Anatomie en Morfologie Wortel De Plantencel Bouw van de plant in zijn functionele delen Vormen waarin die delen zich kunnen voordoen Iets over (geslachtelijke) voortplanting 45 48 Plantencel Stengel Jonge
Anatomie en Morfologie Wortel De Plantencel Bouw van de plant in zijn functionele delen Vormen waarin die delen zich kunnen voordoen Iets over (geslachtelijke) voortplanting 45 48 Plantencel Stengel Jonge
Inhoud les 2. Afwijkingen van wetten van Mendel Polyploïdie. Autopolyploïden Allopolyploïden. les 2 Populatie- en kwantitatieve genetica 1
 Inhoud les 2 Afwijkingen van wetten van Mendel Polyploïdie Autopolyploïden Allopolyploïden genetica 1 Schijnbare afwijkingen van Wetten van Mendel Certatie: letterlijk wedstrijd (tussen pollenkorrels).
Inhoud les 2 Afwijkingen van wetten van Mendel Polyploïdie Autopolyploïden Allopolyploïden genetica 1 Schijnbare afwijkingen van Wetten van Mendel Certatie: letterlijk wedstrijd (tussen pollenkorrels).
Antwoorden Biologie Planten
 Antwoorden Biologie Planten Antwoorden door een scholier 1287 woorden 21 december 2006 6,9 97 keer beoordeeld Vak Methode Biologie Biologie voor jou Opdracht 1 1. Als een deel van een individu uitgroeit
Antwoorden Biologie Planten Antwoorden door een scholier 1287 woorden 21 december 2006 6,9 97 keer beoordeeld Vak Methode Biologie Biologie voor jou Opdracht 1 1. Als een deel van een individu uitgroeit
Samenvattingen. Samenvatting Thema 4: Erfelijkheid. Basisstof 1. Basisstof 2. Erfelijke eigenschappen:
 Samenvatting Thema 4: Erfelijkheid Basisstof 1 Erfelijke eigenschappen: - Genotype: o genen liggen op de chromosomen in kernen van alle cellen o wordt bepaald op moment van de bevruchting - Fenotype: o
Samenvatting Thema 4: Erfelijkheid Basisstof 1 Erfelijke eigenschappen: - Genotype: o genen liggen op de chromosomen in kernen van alle cellen o wordt bepaald op moment van de bevruchting - Fenotype: o
Bestuivingsinformatie
 Bestuivingsinformatie Aardbei Fragaria vesca Familie Ode: Rosales Familie: Rozenfamilie, Rosace. Geslacht: Frigaria Biotoop en bodem: De aardbei prefereert een zonnige standplaats op vochtige vruchtbare
Bestuivingsinformatie Aardbei Fragaria vesca Familie Ode: Rosales Familie: Rozenfamilie, Rosace. Geslacht: Frigaria Biotoop en bodem: De aardbei prefereert een zonnige standplaats op vochtige vruchtbare
Cursus natuurgids LES 2 : PLANTEN DETERMINEREN
 Cursus natuurgids LES 2 : PLANTEN DETERMINEREN 2. Planten determineren, hoe doe je dat? 2.1. Vegetatieve kenmerken 2.2. Voortplantingskenmerken 2.3. APG-indeling en praktische benadering 2.4. Tot slot
Cursus natuurgids LES 2 : PLANTEN DETERMINEREN 2. Planten determineren, hoe doe je dat? 2.1. Vegetatieve kenmerken 2.2. Voortplantingskenmerken 2.3. APG-indeling en praktische benadering 2.4. Tot slot
LES 2 : PLANTEN DETERMINEREN
 Cursus natuurgids LES 2 : PLANTEN DETERMINEREN 2. Planten determineren, hoe doe je dat? 2.1. Vegetatieve kenmerken 2.2. Voortplantingskenmerken 2.3. APG-indeling en praktische benadering 2.4. Tot slot
Cursus natuurgids LES 2 : PLANTEN DETERMINEREN 2. Planten determineren, hoe doe je dat? 2.1. Vegetatieve kenmerken 2.2. Voortplantingskenmerken 2.3. APG-indeling en praktische benadering 2.4. Tot slot
Kleine schorseneer aan het infuus voortgang herstelplan in Drenthe
 Kleine schorseneer aan het infuus voortgang herstelplan in Drenthe WFD-dag, 28 februari 2015 Edwin Dijkhuis Dit project wordt uitgevoerd en mogelijk gemaakt door: Foto: Peter Meininger Portret: Kleine
Kleine schorseneer aan het infuus voortgang herstelplan in Drenthe WFD-dag, 28 februari 2015 Edwin Dijkhuis Dit project wordt uitgevoerd en mogelijk gemaakt door: Foto: Peter Meininger Portret: Kleine
PLANTEN. Basis maakt de vragen 1 t/m 35. Voor iedere vraag kan 1 punt behaald worden
 BK402: PLANTEN Basis maakt de vragen 1 t/m 35. Voor iedere vraag kan 1 punt behaald worden Kader maakt de vragen 1 t/m 45. Voor iedere vraag kan 1 punt behaald worden Beantwoord de volgende vragen. 1 Een
BK402: PLANTEN Basis maakt de vragen 1 t/m 35. Voor iedere vraag kan 1 punt behaald worden Kader maakt de vragen 1 t/m 45. Voor iedere vraag kan 1 punt behaald worden Beantwoord de volgende vragen. 1 Een
Samenvatting. Samenvatting
 Samenvatting Samenvatting 151 Inleiding Samenvatting De meeste eukaryote meercellige organismen planten zich seksueel voort. Dit betekent dat ze gameten produceren, bijvoorbeeld eicellen en sperma, via
Samenvatting Samenvatting 151 Inleiding Samenvatting De meeste eukaryote meercellige organismen planten zich seksueel voort. Dit betekent dat ze gameten produceren, bijvoorbeeld eicellen en sperma, via
Docent: A. Sewsahai KLASSE: 6 VWO
 Henry N. Hassankhan Scholengemeenschap Scholen Gemeenschap Lelydorp [HHS-SGL ARTHUR A. HOOGENDOORN ATHENEUM - VRIJE ATHENEUM AAHA Docent: A. Sewsahai KLASSE: 6 VWO Legenda leerstofafbakening: PAARS: OUDE
Henry N. Hassankhan Scholengemeenschap Scholen Gemeenschap Lelydorp [HHS-SGL ARTHUR A. HOOGENDOORN ATHENEUM - VRIJE ATHENEUM AAHA Docent: A. Sewsahai KLASSE: 6 VWO Legenda leerstofafbakening: PAARS: OUDE
Voortplanting bij planten
 Voortplanting bij planten Opdracht 1 1. Wanneer spreken we van ongeslachtelijke voortplanting? 2. Een uitloper en een wortelstok zijn beide stengels waaraan jonge planten ontstaan. Wat is het verschil
Voortplanting bij planten Opdracht 1 1. Wanneer spreken we van ongeslachtelijke voortplanting? 2. Een uitloper en een wortelstok zijn beide stengels waaraan jonge planten ontstaan. Wat is het verschil
HAVO 5 Begrippenlijst Erfelijkheid allel Allelen zijn verschillende vormen van een gen. Zij liggen in homologe chromosomen op precies dezelfde
 HAVO 5 Begrippenlijst Erfelijkheid allel Allelen zijn verschillende vormen van een gen. Zij liggen in homologe chromosomen op precies dezelfde plaats. Allelen coderen voor dezelfde eigenschap bijvoorbeeld
HAVO 5 Begrippenlijst Erfelijkheid allel Allelen zijn verschillende vormen van een gen. Zij liggen in homologe chromosomen op precies dezelfde plaats. Allelen coderen voor dezelfde eigenschap bijvoorbeeld
I] LAPHE SCALARIS ONTMOET ELAPHE GUTTATA
![I] LAPHE SCALARIS ONTMOET ELAPHE GUTTATA](/thumbs/54/33607375.jpg "I] LAPHE SCALARIS ONTMOET ELAPHE GUTTATA") I] LAPHE SCALARIS ONTMOET ELAPHE GUTTATA EEN UITZONDERLIJK GEVAL VAN HYBRIDISATIE Dr. Emanuele Omattt Via Va/terra 7, 1-4013S Bologna. Italië. Tel. +39 OS1 6441492. Email: tov9602@iperbole.bo/ogna.it Foto's:
I] LAPHE SCALARIS ONTMOET ELAPHE GUTTATA EEN UITZONDERLIJK GEVAL VAN HYBRIDISATIE Dr. Emanuele Omattt Via Va/terra 7, 1-4013S Bologna. Italië. Tel. +39 OS1 6441492. Email: tov9602@iperbole.bo/ogna.it Foto's:
1. Status. Groenknolorchis (Liparis loeselii) H Kenschets. 3. Ecologische vereisten. 4. Huidig voorkomen
 H Kenschets. 3. Ecologische vereisten. 4. Huidig voorkomen") Dit profiel dient gelezen, geïnterpreteerd en gebruikt te worden in combinatie met de leeswijzer, waarin de noodzakelijke uitleg van de verschillende paragrafen vermeld is. Groenknolorchis (Liparis loeselii)
Dit profiel dient gelezen, geïnterpreteerd en gebruikt te worden in combinatie met de leeswijzer, waarin de noodzakelijke uitleg van de verschillende paragrafen vermeld is. Groenknolorchis (Liparis loeselii)
hoofdjes, dat langer is dan deze (Rijksherbarium, Leiden) Bij het gereedmaken van het verspreidingskaartje van Filago pyramidata L.
 Bij het gereedmaken van het verspreidingskaartje van Filago pyramidata L.") De Filago vulgaris-groep in Nederland door F. Adema (Rijksherbarium, Leiden) Bij het gereedmaken van het verspreidingskaartje van Filago pyramidata L. voor de Atlas van de Nederlandse Flora (MENNEMA, 1976)
De Filago vulgaris-groep in Nederland door F. Adema (Rijksherbarium, Leiden) Bij het gereedmaken van het verspreidingskaartje van Filago pyramidata L. voor de Atlas van de Nederlandse Flora (MENNEMA, 1976)
De zaak Parnassia in Drenthe
 De zaak Parnassia in Drenthe risico s van hulp bij lange afstandsverspreiding Gerard Oostermeijer Annie Vos Ben Hoentjen Hans Dekker Karin Uilhoorn Loekie van Tweel Rob Bregman Harry Offringa Willem van
De zaak Parnassia in Drenthe risico s van hulp bij lange afstandsverspreiding Gerard Oostermeijer Annie Vos Ben Hoentjen Hans Dekker Karin Uilhoorn Loekie van Tweel Rob Bregman Harry Offringa Willem van
Insectenbloemen worden dus alleen door bijen bezocht. Hieronder zie je een cartoon waarin beide soorten bloemen zijn afgebeeld.
 Les 3: de bij en de bloem (deel 1) De vorige lessen heb je veel geleerd over de bouw van de bij. Zo heb je goed naar zijn kop en poten gekeken. Door het tekenen weet je nu hoe de bij in elkaar zit en je
Les 3: de bij en de bloem (deel 1) De vorige lessen heb je veel geleerd over de bouw van de bij. Zo heb je goed naar zijn kop en poten gekeken. Door het tekenen weet je nu hoe de bij in elkaar zit en je
Monitoren van klein zeegras, Oosterschelde, september 2008
 Monitoren van klein zeegras, Oosterschelde, 25-26 september 2008 - Wim Giesen, 2 oktober 2008 25-26 september is een bezoek gebracht aan de mitigatielocaties op Tholen (Dortsman Noord en Krabbenkreek Zuid),
Monitoren van klein zeegras, Oosterschelde, 25-26 september 2008 - Wim Giesen, 2 oktober 2008 25-26 september is een bezoek gebracht aan de mitigatielocaties op Tholen (Dortsman Noord en Krabbenkreek Zuid),
Het onderste deel van de stamper is het vruchtbeginsel. Hierin liggen de eicellen. Na bevruchting groeien hier vruchten.
 Bloemen en zaad Voor voortplanting heb je zaad nodig. Maar waar komt zaad vandaan? Om dat te kunnen uitleggen, moet je weten hoe een bloem is opgebouwd en wat bestuiving en bevruchting is. Opbouw van een
Bloemen en zaad Voor voortplanting heb je zaad nodig. Maar waar komt zaad vandaan? Om dat te kunnen uitleggen, moet je weten hoe een bloem is opgebouwd en wat bestuiving en bevruchting is. Opbouw van een
Voorbereiding toelatingsexamen arts/tandarts. Biologie: evolutieleer 6/29/2013. dr. Brenda Casteleyn
 Voorbereiding toelatingsexamen arts/tandarts Biologie: evolutieleer 6/29/2013 dr. Brenda Casteleyn Met dank aan: Leen Goyens (http://users.telenet.be/toelating) en studenten van forum http://www.toelatingsexamen-geneeskunde.be
Voorbereiding toelatingsexamen arts/tandarts Biologie: evolutieleer 6/29/2013 dr. Brenda Casteleyn Met dank aan: Leen Goyens (http://users.telenet.be/toelating) en studenten van forum http://www.toelatingsexamen-geneeskunde.be
Antwoorden door een scholier 1825 woorden 28 februari keer beoordeeld
 Antwoorden door een scholier 1825 woorden 28 februari 2005 6 511 keer beoordeeld Vak Biologie Biologie Thema 5 Opdracht 1 1. Het uiterlijk is een fenotype van een organisme. 2. Alle erfelijke informatie
Antwoorden door een scholier 1825 woorden 28 februari 2005 6 511 keer beoordeeld Vak Biologie Biologie Thema 5 Opdracht 1 1. Het uiterlijk is een fenotype van een organisme. 2. Alle erfelijke informatie
Hyngsteblom, Knineblêdden en Tiksel
 Hyngsteblom, Knineblêdden en Tiksel Piet Oosterveld (IBN-DLO, Postbus 23, 6700 AA Wageningen) On Frisian dandelions Some relatively rare Taraxacum species of the sections Palustria and Celtica occurring
Hyngsteblom, Knineblêdden en Tiksel Piet Oosterveld (IBN-DLO, Postbus 23, 6700 AA Wageningen) On Frisian dandelions Some relatively rare Taraxacum species of the sections Palustria and Celtica occurring
4 HAVO thema 4 Erfelijkheid EXAMENTRAINER OEFENVRAGEN
 Examentrainer Vragen Karyogrammen In afbeelding 1 zijn twee karyogrammen weergegeven. Deze karyogrammen zijn afkomstig van een eeneiige tweeling. Het ene kind is van het mannelijk geslacht zonder duidelijke
Examentrainer Vragen Karyogrammen In afbeelding 1 zijn twee karyogrammen weergegeven. Deze karyogrammen zijn afkomstig van een eeneiige tweeling. Het ene kind is van het mannelijk geslacht zonder duidelijke
6,1. Samenvatting door een scholier 1949 woorden 7 februari keer beoordeeld. Biologie voor jou
 Samenvatting door een scholier 1949 woorden 7 februari 2011 6,1 46 keer beoordeeld Vak Methode Biologie Biologie voor jou Basisstof 1: chromosomen Chromosomen: komen voor in een celkern bevindt zich de
Samenvatting door een scholier 1949 woorden 7 februari 2011 6,1 46 keer beoordeeld Vak Methode Biologie Biologie voor jou Basisstof 1: chromosomen Chromosomen: komen voor in een celkern bevindt zich de
CGM/ Advies: Ontwikkelingsstadia en levensvatbaarheid hybride planten
 BEZOEKADRES: A. VAN LEEUWENHOEKLAAN 9 3721 MA BILTHOVEN POSTADRES: POSTBUS 578 3720 AN BILTHOVEN Aan de minister van Volkshuisvesting, Ruimtelijke Ordening en Milieubeheer Mevrouw J.C. Huizinga-Heringa
BEZOEKADRES: A. VAN LEEUWENHOEKLAAN 9 3721 MA BILTHOVEN POSTADRES: POSTBUS 578 3720 AN BILTHOVEN Aan de minister van Volkshuisvesting, Ruimtelijke Ordening en Milieubeheer Mevrouw J.C. Huizinga-Heringa
Anacamptis pyramidalis bij Wijk aan Zee. door. de Vries-laboratorium, Amsterdam) worden de tellingen grafisch weergegeven.
 worden de tellingen grafisch weergegeven.") Anacamptis pyramidalis bij Wijk aan Zee door A.A. Sterk (Hugo de Vries-laboratorium, Amsterdam) In Gorteria schrijft DE WILDE (1976) een artikel over de verarming van de duinflora bij Wijk aan Zee. De
Anacamptis pyramidalis bij Wijk aan Zee door A.A. Sterk (Hugo de Vries-laboratorium, Amsterdam) In Gorteria schrijft DE WILDE (1976) een artikel over de verarming van de duinflora bij Wijk aan Zee. De
soortscomplexen het verschijnsel apomixis geaccentueerd amphimictische voortplanting apoamphimicten. planten met zowel apoals
 Enige opmerkingen over Hieracium pilosella L. in Nederland door Th.W.J. Gadella en E. Kliphuis (Instituut voor Systematische Plantkunde, Utrecht) Inleiding Het een zeer groot aantal soorten en ondersoorten
Enige opmerkingen over Hieracium pilosella L. in Nederland door Th.W.J. Gadella en E. Kliphuis (Instituut voor Systematische Plantkunde, Utrecht) Inleiding Het een zeer groot aantal soorten en ondersoorten
ERFELIJKHEID. 1 N i e t a l l e m a a l h e t z e l f d e Afbeelding 17-2
 ERFELIJKHEID 1 N i e t a l l e m a a l h e t z e l f d e Afbeelding 17-2 Afbeelding 17-1 Mensen uit elkaar houden vind je vast makkelijker. Toch hebben ook mensen veel meer overeenkomsten dan verschillen.
ERFELIJKHEID 1 N i e t a l l e m a a l h e t z e l f d e Afbeelding 17-2 Afbeelding 17-1 Mensen uit elkaar houden vind je vast makkelijker. Toch hebben ook mensen veel meer overeenkomsten dan verschillen.
NIEUWSBRIEF FLORON-FWT NR. 21, APRIL 2000
 NIEUWSBRIEF FLORON-FWT NR. 21, APRIL 2000 Inleiding Paardenbloemen in Twente (1) O.G. Zijlstra De afgelopen tien jaar is er in Twente, vooral in en rond Hengelo en Enschede, een begin gemaakt met de inventarisatie
NIEUWSBRIEF FLORON-FWT NR. 21, APRIL 2000 Inleiding Paardenbloemen in Twente (1) O.G. Zijlstra De afgelopen tien jaar is er in Twente, vooral in en rond Hengelo en Enschede, een begin gemaakt met de inventarisatie
Toerisme in perspectief. NBTC Holland Marketing Afdeling Research
 Toerisme in perspectief NBTC Holland Marketing Afdeling Research Inleiding In dit rapport wordt op hoofdlijnen een beeld geschetst van trends en ontwikkelingen in het (internationaal) toerisme en de factoren
Toerisme in perspectief NBTC Holland Marketing Afdeling Research Inleiding In dit rapport wordt op hoofdlijnen een beeld geschetst van trends en ontwikkelingen in het (internationaal) toerisme en de factoren
Wespenorchis (Epipactis)
") Wespenorchis (Epipactis) LPW-Florasleutel samengesteld door Felix Baeten ALGEMENE SLEUTEL Onderste lip met smalle insnoering Beweeglijk eindstuk (epichiel), bloem bruin en wit Moeraswespenorchis Geen smalle
Wespenorchis (Epipactis) LPW-Florasleutel samengesteld door Felix Baeten ALGEMENE SLEUTEL Onderste lip met smalle insnoering Beweeglijk eindstuk (epichiel), bloem bruin en wit Moeraswespenorchis Geen smalle
De volgende vragen testen je kennis van de meest voorkomende vaktermen in de klassieke genetica. Welk woord ontbreekt in de volgende zinnen?
 Populatiegenetica Versie 2006-2007 Vragen bij COO Belangrijke begrippen De volgende vragen testen je kennis van de meest voorkomende vaktermen in de klassieke genetica. Welk woord ontbreekt in de volgende
Populatiegenetica Versie 2006-2007 Vragen bij COO Belangrijke begrippen De volgende vragen testen je kennis van de meest voorkomende vaktermen in de klassieke genetica. Welk woord ontbreekt in de volgende
Nederlandse samenvatting
 138 Uitstel van ouderschap De positie van de vrouw in de westerse maatschappij is de laatste tientallen jaren fundamenteel veranderd. Vrouwen zijn hoger opgeleid dan vroeger en werken vaker buitenshuis.
138 Uitstel van ouderschap De positie van de vrouw in de westerse maatschappij is de laatste tientallen jaren fundamenteel veranderd. Vrouwen zijn hoger opgeleid dan vroeger en werken vaker buitenshuis.
Voortplanting en celdeling
 Voortplanting en celdeling Inleiding Kenmerkend voor de levende natuur is het streven om de soort in stand te houden. Voor de groene plant als groep komt daar nog bij het vastleggen en beschikbaar maken
Voortplanting en celdeling Inleiding Kenmerkend voor de levende natuur is het streven om de soort in stand te houden. Voor de groene plant als groep komt daar nog bij het vastleggen en beschikbaar maken
Teveel centrosomen in kankercellen
 Teveel centrosomen in kankercellen Ongecoördineerde celdeling zorgt ervoor dat kankercellen steeds verder ontsporen, denkt een onderzoeksgroep van oncologen uit Boston. Kankercellen zijn vaak aneuploïd:
Teveel centrosomen in kankercellen Ongecoördineerde celdeling zorgt ervoor dat kankercellen steeds verder ontsporen, denkt een onderzoeksgroep van oncologen uit Boston. Kankercellen zijn vaak aneuploïd:
CGM/ Advies Inperkingsmaatregelen voor werkzaamheden met genetisch gemodificeerde Geranium robertianum en Geranium pyrenaicum
 Aan de staatssecretaris van Infrastructuur en Milieu Mevrouw W.J. Mansveld Postbus 20901 2500 EX Den Haag DATUM 18 mei 2015 KENMERK ONDERWERP CGM/150518-01 Advies Inperkingsmaatregelen voor werkzaamheden
Aan de staatssecretaris van Infrastructuur en Milieu Mevrouw W.J. Mansveld Postbus 20901 2500 EX Den Haag DATUM 18 mei 2015 KENMERK ONDERWERP CGM/150518-01 Advies Inperkingsmaatregelen voor werkzaamheden
Bijen en bestuiving in de fruitteelt bij open teelten
 Bijen en bestuiving in de fruitteelt bij open teelten ABTB ANI LLTB ZLTO VBBN Bijen en bestuiving van fruit bij open teelten Bloei, bestuiving en bevruchting spelen een essentiële rol bij de vorming van
Bijen en bestuiving in de fruitteelt bij open teelten ABTB ANI LLTB ZLTO VBBN Bijen en bestuiving van fruit bij open teelten Bloei, bestuiving en bevruchting spelen een essentiële rol bij de vorming van
Door recombinatie ontstaat een grote vescheidenheid in genotypen binnen een soort. (genetische
 Chromosomen bestaan voor een groot deel uit DNA DNA bevat de erfelijke informatie van een organisme. Een gen(ook wel erffactor) is een stukje DNA dat de informatie bevat voor een erfelijke eigenschap(bvb
Chromosomen bestaan voor een groot deel uit DNA DNA bevat de erfelijke informatie van een organisme. Een gen(ook wel erffactor) is een stukje DNA dat de informatie bevat voor een erfelijke eigenschap(bvb
6,4. Samenvatting door E woorden 6 december keer beoordeeld. Biologie voor jou
 Samenvatting door E. 1393 woorden 6 december 2016 6,4 18 keer beoordeeld Vak Methode Biologie Biologie voor jou Thema 4: Erfelijkheid 5-HTTPLR gen heeft invloed op de hoeveelheid geluk die je ervaart.
Samenvatting door E. 1393 woorden 6 december 2016 6,4 18 keer beoordeeld Vak Methode Biologie Biologie voor jou Thema 4: Erfelijkheid 5-HTTPLR gen heeft invloed op de hoeveelheid geluk die je ervaart.
Juli blauw Biologie Vraag 1
 Biologie Vraag 1 Bij bijen komt parthenogenese voor. Dit is de ontwikkeling van een individu uit een onbevruchte eicel. Bij bijen ontstaan de darren (mannelijke bijen) parthenogenetisch. De koningin en
Biologie Vraag 1 Bij bijen komt parthenogenese voor. Dit is de ontwikkeling van een individu uit een onbevruchte eicel. Bij bijen ontstaan de darren (mannelijke bijen) parthenogenetisch. De koningin en
Juli geel Biologie Vraag 1
 Biologie Vraag 1 Bij bijen komt parthenogenese voor. Dit is de ontwikkeling van een individu uit een onbevruchte eicel. Bij bijen ontstaan de darren (mannelijke bijen) parthenogenetisch. De koningin en
Biologie Vraag 1 Bij bijen komt parthenogenese voor. Dit is de ontwikkeling van een individu uit een onbevruchte eicel. Bij bijen ontstaan de darren (mannelijke bijen) parthenogenetisch. De koningin en
Voortplanting. Lesbrief. Werkgroep Schoolactiviteiten. I.V.N. afd.hengelo. Tel. O74 2770390
 Voortplanting Lesbrief Werkgroep Schoolactiviteiten I.V.N. afd.hengelo Tel. O74 2770390 1 Deze lesbrief wordt U aangeboden door het I.V.N. afd. Hengelo Voortplanting = zorgen voor jonge planten A. Inleiding
Voortplanting Lesbrief Werkgroep Schoolactiviteiten I.V.N. afd.hengelo Tel. O74 2770390 1 Deze lesbrief wordt U aangeboden door het I.V.N. afd. Hengelo Voortplanting = zorgen voor jonge planten A. Inleiding
Inheemse plantensoorten versus exoten. Stellingen. Definities 08/02/2012. Workshop Inheemse plantensoorten versus exoten, Marco Hoffman, PPO
 Workshop Groendag Hoorn Inheemse plantensoorten versus exoten Marco Hoffman Opzet workshop Peilen meningen deelnemers & enkele definities Achtergronden brochure Feiten en objectieve achtergrondinformatie
Workshop Groendag Hoorn Inheemse plantensoorten versus exoten Marco Hoffman Opzet workshop Peilen meningen deelnemers & enkele definities Achtergronden brochure Feiten en objectieve achtergrondinformatie
Boterbloem (Ranunculus)
") Boterbloem (Ranunculus) LPW-Florasleutel samengesteld door Johan Geusens ALGEMENE SLEUTEL Boterbloemen van natte, vochtige plaatsen : Grote boterbloem Blaartrekkende boterbloem Egelboterbloem Boterbloemen
Boterbloem (Ranunculus) LPW-Florasleutel samengesteld door Johan Geusens ALGEMENE SLEUTEL Boterbloemen van natte, vochtige plaatsen : Grote boterbloem Blaartrekkende boterbloem Egelboterbloem Boterbloemen
Karakterisatie van stammen van de aardappelziekte in Wallonië (2014)
") Karakterisatie van stammen van de aardappelziekte in Wallonië (2014) V. César (CRA-W) Samenvatting Het Waals onderzoekscentrum voor de landbouw onderzoekt sinds 1999 de populaties van de aardappelplaag.
Karakterisatie van stammen van de aardappelziekte in Wallonië (2014) V. César (CRA-W) Samenvatting Het Waals onderzoekscentrum voor de landbouw onderzoekt sinds 1999 de populaties van de aardappelplaag.
Nederlandse samenvatting
 Nederlandse samenvatting Tot voor kort werd de relatie tussen bloemplanten en hun bestuivers vooral gezien als gespecialiseerd en wederzijds sterk aangepast. Specialisatie betekent in dit geval dat één
Nederlandse samenvatting Tot voor kort werd de relatie tussen bloemplanten en hun bestuivers vooral gezien als gespecialiseerd en wederzijds sterk aangepast. Specialisatie betekent in dit geval dat één
d rm Neder wa e landopg
 Opgewarmd Nederland deel Plant en dier: blijven, komen, weggaan of... Soorten, verspreiding en klimaat Kleine beestjes: sterk in beweging Libellen: voordeel van een warmer klimaat Dagvlinders: extra onder
Opgewarmd Nederland deel Plant en dier: blijven, komen, weggaan of... Soorten, verspreiding en klimaat Kleine beestjes: sterk in beweging Libellen: voordeel van een warmer klimaat Dagvlinders: extra onder
Inleiding. Seizoensveranderingen bij bladeren van Taraxacum. (Afd. Plantensystematiek, Biologisch Laboratorium, Vrije Universiteit, Amsterdam)
") Seizoensveranderingen bij bladeren van Taraxacum door J.H. Ietswaart en M. Vroman (Afd. Plantensystematiek, Biologisch Laboratorium, Vrije Universiteit, Amsterdam) Inleiding Het is bekend dat bij Taraxacum-
Seizoensveranderingen bij bladeren van Taraxacum door J.H. Ietswaart en M. Vroman (Afd. Plantensystematiek, Biologisch Laboratorium, Vrije Universiteit, Amsterdam) Inleiding Het is bekend dat bij Taraxacum-
Basterdwederik (Epilobium)
") ALGEMENE SLEUTEL Basterdwederik (Epilobium) LPW-Florasleutel samengesteld door Bert Berten Bladen verspreid Gewoon wilgenroosje Bladen tegenoverstaand; stempel in kruis of knotsvormig Stempel in 4 gespleten,
ALGEMENE SLEUTEL Basterdwederik (Epilobium) LPW-Florasleutel samengesteld door Bert Berten Bladen verspreid Gewoon wilgenroosje Bladen tegenoverstaand; stempel in kruis of knotsvormig Stempel in 4 gespleten,
Paardenbloemen in Twente (1)
") Paardenbloemen in Twente (1) Inleiding O.G. Zijlstra De afgelopen tien jaar is er in Twente, vooral in en rond Hengelo en Enschede, een begin gemaakt met de inventarisatie van de recente paardenbloemenflora.
Paardenbloemen in Twente (1) Inleiding O.G. Zijlstra De afgelopen tien jaar is er in Twente, vooral in en rond Hengelo en Enschede, een begin gemaakt met de inventarisatie van de recente paardenbloemenflora.
De Wiershoeck-Kinderwerktuin, dinsdag 21 maart Beste natuurliefhebber/-ster,
 De Wiershoeck-Kinderwerktuin, dinsdag 21 maart 2017 Beste natuurliefhebber/-ster, Het was dinsdagmorgen mooi zonnig weer, maar er stond een stevige wind. Voor de middag heeft het nog even gehageld. Het
De Wiershoeck-Kinderwerktuin, dinsdag 21 maart 2017 Beste natuurliefhebber/-ster, Het was dinsdagmorgen mooi zonnig weer, maar er stond een stevige wind. Voor de middag heeft het nog even gehageld. Het
Academisch cafe Utrechts Universiteitsfonds, 12 april 2007, Arnhem
 Academisch cafe Utrechts Universiteitsfonds, 12 april 2007, Arnhem Geachte aanwezigen, mijn naam is Roy Erkens en de komende 15 minuten ga ik u iets vertellen over mijn proefschrift getiteld from morphological
Academisch cafe Utrechts Universiteitsfonds, 12 april 2007, Arnhem Geachte aanwezigen, mijn naam is Roy Erkens en de komende 15 minuten ga ik u iets vertellen over mijn proefschrift getiteld from morphological
Monitoren van klein zeegras, Oosterschelde, augustus 2008
 Monitoren van klein zeegras, Oosterschelde, 26-28 augustus 2008 - Wim Giesen, 30 augustus 2008 25-28 augustus 2008 is een bezoek gebracht aan de mitigatielocaties op Tholen (Dortsman Noord en Krabbenkreek
Monitoren van klein zeegras, Oosterschelde, 26-28 augustus 2008 - Wim Giesen, 30 augustus 2008 25-28 augustus 2008 is een bezoek gebracht aan de mitigatielocaties op Tholen (Dortsman Noord en Krabbenkreek
Aponogeton rara ' Door : Liesbeth van Bruggen
 WWW.WATERPLANTEN.ORG Aponogeton rara ' Door : Liesbeth van Bruggen Dit verhaal gaat niet over een nieuw-ontdekte Aponogeton. Het gaat ook niet over een zeldzame Aponogeton (dan had het verhaal 'Aponogeton
WWW.WATERPLANTEN.ORG Aponogeton rara ' Door : Liesbeth van Bruggen Dit verhaal gaat niet over een nieuw-ontdekte Aponogeton. Het gaat ook niet over een zeldzame Aponogeton (dan had het verhaal 'Aponogeton
Biologie (jaartal onbekend)
") Biologie (jaartal onbekend) 1) Bijgevoegde fotografische afbeelding geeft de elektronenmicroscopische opname van een organel (P) van een cel. Wat is de belangrijkste functie van dit organel? A. Het transporteren
Biologie (jaartal onbekend) 1) Bijgevoegde fotografische afbeelding geeft de elektronenmicroscopische opname van een organel (P) van een cel. Wat is de belangrijkste functie van dit organel? A. Het transporteren
Aan de minister van Volkshuisvesting, Ruimtelijke Ordening en Milieubeheer Mevrouw dr. J.M. Cramer Postbus GX Den Haag
 Aan de minister van Volkshuisvesting, Ruimtelijke Ordening en Milieubeheer Mevrouw dr. J.M. Cramer Postbus 30945 2500 GX Den Haag DATUM 26 november 2007 KENMERK CGM/071126-02 ONDERWERP Advies inschaling
Aan de minister van Volkshuisvesting, Ruimtelijke Ordening en Milieubeheer Mevrouw dr. J.M. Cramer Postbus 30945 2500 GX Den Haag DATUM 26 november 2007 KENMERK CGM/071126-02 ONDERWERP Advies inschaling
2. Erfelijkheid en de ziekte van Huntington
 2. Erfelijkheid en de ziekte van Huntington Erfelijkheid Erfelijk materiaal in de 46 chromosomen De mens heeft in de kern van elke lichaamscel 46 chromosomen: het gaat om 22 paar lichaamsbepalende chromosomen
2. Erfelijkheid en de ziekte van Huntington Erfelijkheid Erfelijk materiaal in de 46 chromosomen De mens heeft in de kern van elke lichaamscel 46 chromosomen: het gaat om 22 paar lichaamsbepalende chromosomen
populaties komen beide geslachten door elkaar voor, zodat vruchtvorming Onvruchtbaarheid, een vruchtbaar terrein van studie
 Onvruchtbaarheid, een vruchtbaar terrein van studie door T.W.J. Gadella (Instituut voor Systematische Plantkunde, Utrecht) JACOBS (1969, 1970) vestigt in een tweetal tijdschriftartikelen de aandacht op
Onvruchtbaarheid, een vruchtbaar terrein van studie door T.W.J. Gadella (Instituut voor Systematische Plantkunde, Utrecht) JACOBS (1969, 1970) vestigt in een tweetal tijdschriftartikelen de aandacht op
Download het originele werkstuk in de bijlage voor de foto's van de bloemen.
 Praktische-opdracht door een scholier 1556 woorden 24 augustus 2010 5,9 20 keer beoordeeld Vak Biologie Download het originele werkstuk in de bijlage voor de foto's van de bloemen. Inhoudsopgave Inleiding
Praktische-opdracht door een scholier 1556 woorden 24 augustus 2010 5,9 20 keer beoordeeld Vak Biologie Download het originele werkstuk in de bijlage voor de foto's van de bloemen. Inhoudsopgave Inleiding
Aan de Minister van Volkshuisvesting, Ruimtelijke Ordening en Milieubeheer Mevrouw dr. J.M. Cramer Postbus GX Den Haag
 Aan de Minister van Volkshuisvesting, Ruimtelijke Ordening en Milieubeheer Mevrouw dr. J.M. Cramer Postbus 30945 2500 GX Den Haag DATUM 23 juni 2008 KENMERK CGM/080623-02 ONDERWERP Inschaling van handelingen
Aan de Minister van Volkshuisvesting, Ruimtelijke Ordening en Milieubeheer Mevrouw dr. J.M. Cramer Postbus 30945 2500 GX Den Haag DATUM 23 juni 2008 KENMERK CGM/080623-02 ONDERWERP Inschaling van handelingen
Toerisme in perspectief
 Toerisme in perspectief NBTC afdeling Research Leidschendam 10-7-2012 Inleiding In dit rapport wordt op hoofdlijnen een beeld geschetst van trends en ontwikkelingen in het (internationaal) toerisme en
Toerisme in perspectief NBTC afdeling Research Leidschendam 10-7-2012 Inleiding In dit rapport wordt op hoofdlijnen een beeld geschetst van trends en ontwikkelingen in het (internationaal) toerisme en
A bat friendly colour spectrum? Effecten van klimaatverandering op vleermuizen
 A bat friendly colour spectrum? Effecten van klimaatverandering op vleermuizen Herman Limpens en Jasja Dekker Effecten van klimaatverandering op vleermuizen?? Vooral: analyserende speculerende verhalen
A bat friendly colour spectrum? Effecten van klimaatverandering op vleermuizen Herman Limpens en Jasja Dekker Effecten van klimaatverandering op vleermuizen?? Vooral: analyserende speculerende verhalen
Aantal huisartsen en aantal FTE van huisartsen vanaf 2007 tot en met 2016
 Aantal huisartsen en aantal FTE van huisartsen vanaf 2007 tot en met 2016 Werken er nu meer of minder huisartsen dan 10 jaar geleden en werken zij nu meer of minder FTE? LF.J. van der Velden & R.S. Batenburg,
Aantal huisartsen en aantal FTE van huisartsen vanaf 2007 tot en met 2016 Werken er nu meer of minder huisartsen dan 10 jaar geleden en werken zij nu meer of minder FTE? LF.J. van der Velden & R.S. Batenburg,
Leni Duistermaat, Naturalis sectie Botanie, onderzoeker Nederlandse flora
 Herbariummateriaal verzamelen nog van deze tijd?! Leni Duistermaat, Naturalis sectie Botanie, onderzoeker Nederlandse flora Leni.Duistermaat@naturalis.nl Nederland kent een lange traditie van het inventariseren
Herbariummateriaal verzamelen nog van deze tijd?! Leni Duistermaat, Naturalis sectie Botanie, onderzoeker Nederlandse flora Leni.Duistermaat@naturalis.nl Nederland kent een lange traditie van het inventariseren
HAPTE CHAP SAMENVATTING
 HAPTE CHAP Wanneer voortplanting tussen individuen van verschillende soorten, maar ook van verschillende populaties wordt voorkómen, noemen we dit reproductieve isolatie. Reproductieve isolatie speelt
HAPTE CHAP Wanneer voortplanting tussen individuen van verschillende soorten, maar ook van verschillende populaties wordt voorkómen, noemen we dit reproductieve isolatie. Reproductieve isolatie speelt
Ongeslachtelijke voortplanting vmbo-b34
 Auteur Laatst gewijzigd Licentie Webadres VO-content 12 juli 2016 CC Naamsvermelding 3.0 Nederland licentie https://maken.wikiwijs.nl/73623 Dit lesmateriaal is gemaakt met Wikiwijs van Kennisnet. Wikiwijs
Auteur Laatst gewijzigd Licentie Webadres VO-content 12 juli 2016 CC Naamsvermelding 3.0 Nederland licentie https://maken.wikiwijs.nl/73623 Dit lesmateriaal is gemaakt met Wikiwijs van Kennisnet. Wikiwijs
Samenvatting Planten VMBO 4a Biologie voor Jou
 Samenvatting Planten VMBO 4a Biologie voor Jou 2.1 Ongeslachtelijke voortplanting = voortplanting waarbij geen bevruchting plaats vindt; hierbij groeit een stukje van de volwassen plant uit tot een nieuwe
Samenvatting Planten VMBO 4a Biologie voor Jou 2.1 Ongeslachtelijke voortplanting = voortplanting waarbij geen bevruchting plaats vindt; hierbij groeit een stukje van de volwassen plant uit tot een nieuwe
Levende Natuur 1989 nummer 4 98
 Levende Natuur 989 nummer 4 98 Foto. T.gelricum in het natuurreservaat Arkemheen (Nekkeveld, gem. Nijkerk). Het reservaat is binnendijks gelegen aan de voet van de dijk van het emmeer. In de onbeweide
Levende Natuur 989 nummer 4 98 Foto. T.gelricum in het natuurreservaat Arkemheen (Nekkeveld, gem. Nijkerk). Het reservaat is binnendijks gelegen aan de voet van de dijk van het emmeer. In de onbeweide
2 Voortplanten met organen Bouw en werking van geslachtsorganen Werking van geslachtshormonen Afsluiting 31
 Inhoud Voorwoord 5 Inleiding 6 1 Voortplanten van genen 9 1.1 Genetica 9 1.2 Kruisingen 13 1.3 Crossing-over en mutatie 16 1.4 Erfelijkheid en praktijk 17 1.5 Inteelt en inteeltdepressie 21 1.6 Afsluiting
Inhoud Voorwoord 5 Inleiding 6 1 Voortplanten van genen 9 1.1 Genetica 9 1.2 Kruisingen 13 1.3 Crossing-over en mutatie 16 1.4 Erfelijkheid en praktijk 17 1.5 Inteelt en inteeltdepressie 21 1.6 Afsluiting
Wijnimport Nederland naar regio
 DO RESEARCH Wijnimport Nederland naar regio Sterke opmars wijn uit Chili Jeroen den Ouden 1-10-2011 Inleiding en inhoudsopgave Pagina I De invoer van wijn in Nederland 1 II De invoer van wijn naar herkomst
DO RESEARCH Wijnimport Nederland naar regio Sterke opmars wijn uit Chili Jeroen den Ouden 1-10-2011 Inleiding en inhoudsopgave Pagina I De invoer van wijn in Nederland 1 II De invoer van wijn naar herkomst
Samenvatting Erfelijkheid Vmbo 3a Biologie voor Jou. Erfelijke informatie ligt in de celkern in de chromosomen. Chromosomen bestaan weer uit DNA.
 Samenvatting Erfelijkheid Vmbo 3a Biologie voor Jou 4.1 Fenotype Genotype = waarneembare eigenschappen van een individu = de erfelijke informatie in het DNA Genotype + milieufactoren = fenotype Erfelijke
Samenvatting Erfelijkheid Vmbo 3a Biologie voor Jou 4.1 Fenotype Genotype = waarneembare eigenschappen van een individu = de erfelijke informatie in het DNA Genotype + milieufactoren = fenotype Erfelijke
3.Mitose. 2.Mitose. Hoeveel chromatiden bevat een menselijke cel maximaal tijdens te mitose?
 In welke cellen vindt mitose plaats? Hoeveel chromatiden bevat een menselijke cel maximaal tijdens te mitose? Hoeveel centromeren bevat een menselijke cel maximaal tijdens de mitose? Een cel ondergaat
In welke cellen vindt mitose plaats? Hoeveel chromatiden bevat een menselijke cel maximaal tijdens te mitose? Hoeveel centromeren bevat een menselijke cel maximaal tijdens de mitose? Een cel ondergaat
De fascinerende wereld van wilde planten Samenvatting van inleiding Jacques de Raad voor de IVN-gidsencursus op 18 april 2011
 De fascinerende wereld van wilde planten Samenvatting van inleiding Jacques de Raad voor de IVN-gidsencursus op 18 april 2011 Om duidelijk te maken waar we het over hebben eerst wat getallen. In Nederland
De fascinerende wereld van wilde planten Samenvatting van inleiding Jacques de Raad voor de IVN-gidsencursus op 18 april 2011 Om duidelijk te maken waar we het over hebben eerst wat getallen. In Nederland
Vervolgens wordt ook ingegaan op de hypothese dat patronen in fenotypische plasticiteit adaptief kunnen zijn. De eerste vraag was of natuurlijke
 Variatie in abiotische factoren komt voor in alle natuurlijke omgevingen, zoals variatie in ruimte en tijd in temperatuur, licht en vochtigheid. Deze factoren kunnen vervolgens bijvoorbeeld voedselaanbod
Variatie in abiotische factoren komt voor in alle natuurlijke omgevingen, zoals variatie in ruimte en tijd in temperatuur, licht en vochtigheid. Deze factoren kunnen vervolgens bijvoorbeeld voedselaanbod
2 Bemesting 44 2.1 Meststoffen 44 2.2 Soorten meststoffen 46 2.3 Grondonderzoek 49 2.4 Mestwetgeving 49
 Inhoud Voorwoord 5 Inleiding 6 1 Bodem en grond 9 1.1 Grond, bodem en grondsoorten 9 1.2 Eigenschappen van grond 20 1.3 Problemen met de grond 23 1.4 Verbeteren van landbouwgronden 30 1.5 Transport van
Inhoud Voorwoord 5 Inleiding 6 1 Bodem en grond 9 1.1 Grond, bodem en grondsoorten 9 1.2 Eigenschappen van grond 20 1.3 Problemen met de grond 23 1.4 Verbeteren van landbouwgronden 30 1.5 Transport van
Samenvatting Biologie Planten en cellen
 Samenvatting Biologie Planten en cellen Samenvatting door een scholier 1333 woorden 5 juni 2004 5,6 147 keer beoordeeld Vak Biologie Planten Planten zijn overal om ons heen. Bomen en struiken. De een opvallend
Samenvatting Biologie Planten en cellen Samenvatting door een scholier 1333 woorden 5 juni 2004 5,6 147 keer beoordeeld Vak Biologie Planten Planten zijn overal om ons heen. Bomen en struiken. De een opvallend
Thema 3b Voortplanting. Wereldoriëntatie L6
 Wereldoriëntatie L6 Les 1 Instandhouding van de soort. p. 3 1. Alle levende wezens zijn opgebouwd uit cellen. Vul de woorden celkern, celvloeistof en celwand correct aan. 2. Vul de tekst aan. In elke
Wereldoriëntatie L6 Les 1 Instandhouding van de soort. p. 3 1. Alle levende wezens zijn opgebouwd uit cellen. Vul de woorden celkern, celvloeistof en celwand correct aan. 2. Vul de tekst aan. In elke
Cover Page. The handle holds various files of this Leiden University dissertation
 Cover Page The handle http://hdl.handle.net/1887/45834 holds various files of this Leiden University dissertation Author: Evans, Tracy Title: Management implications for invertebrate assemblages in the
Cover Page The handle http://hdl.handle.net/1887/45834 holds various files of this Leiden University dissertation Author: Evans, Tracy Title: Management implications for invertebrate assemblages in the
Lang bloeiende inheemse vaste planten voor bijen en vlinders
 www.bloembergecologie.com Lang bloeiende inheemse vaste planten voor bijen en vlinders Bakkers, Sanne; Bloemberg, Mark HAS Hogeschool s-hertogenbosch, 26 juni 2015 Bloemberg M.S. 07-05-2017 1 Lang bloeiende
www.bloembergecologie.com Lang bloeiende inheemse vaste planten voor bijen en vlinders Bakkers, Sanne; Bloemberg, Mark HAS Hogeschool s-hertogenbosch, 26 juni 2015 Bloemberg M.S. 07-05-2017 1 Lang bloeiende
PLANTEN VOORTPLANTING
 . PLANTEN VOORTPLANTING Voortplanting - Ongeslachtelijke voortplanting nakomelingen komen van 1 ouderplant * natuurlijke manieren * kunstmatige manieren - Geslachtelijke voortplanting uitwisseling van
. PLANTEN VOORTPLANTING Voortplanting - Ongeslachtelijke voortplanting nakomelingen komen van 1 ouderplant * natuurlijke manieren * kunstmatige manieren - Geslachtelijke voortplanting uitwisseling van