Ontwikkeling van het bodemleven in de vooroever na aanleg van een onderwatersuppletie. Case studie Ameland en Schiermonnikoog
|
|
|
- Diana de Smedt
- 5 jaren geleden
- Aantal bezoeken:
Transcriptie

1 Ontwikkeling van het bodemleven in de vooroever na aanleg van een onderwatersuppletie Case
2
3 Ontwikkeling van het bodemleven in de vooroever na aanleg van een onderwatersuppletie Case S.A. Vergouwen MSc ir. H. Meijer - Holzhauer Deltares, 2016, B
4


5 Deltares Titel Ontwikkeling van het bodemleven in de vooroever na aanleg van een onderwatersuppletie Opdrachtgever Rijkswaterstaat Project Kenmerk Pagina's ZKS Trefwoorden Suppletierichtlijn Ameland, kustlijn, macrobenthos, onderwatersuppletie, vooroever Samenvatting Hoewel zandsuppleties worden gezien als noodzakelijk voor het handhaven van de functies van de kust, bestaat er de zorg dat de huidige zandsuppleties van invloed zijn op het functioneren van het ecosysteem van de ondiepe kustzone. Daarom is door RWS in 2009 een samenwerkingsovereenkomst opgesteld met vier natuurbeschermingsorganisaties waarin is afgesproken om van 2009 tim 2015 onderzoek te doen naar mogelijke effecten van zandsuppleties op het kustecosysteem. In dit kader is er in 2009 gekozen om de ontwikkeling van het bodemleven na de zandsuppletie op Ameland-midden in te volgen voor een periode van vier jaar en daarmee een volledige suppletiecyclus te beslaan. Het onderzoek is uitgevoerd binnen het kader van programma Kennis voor Primaire Processen Beheer & Onderhoud Kust. Dit rapport geeft een overzicht van de ontwikkelingen van de bodemdieren van de vooroever op Ameland tussen Tijdens deze studie zijn er op Ameland twee deelgebieden onderzocht; Ameland Impact, waar in 2011 is gesuppleerd, en Ameland Uitstraling, een gebied direct ten oosten van de suppletie, dat mogelijk indirecte effecten kan ondervinden van de suppletie. Schiermonnikoog is daarnaast door de tijd gevolgd als referentiegebied. In deze studie zijn metereologische en sedimentsamenstelling meegenomen, om inzicht te geven in de fysische omstandigheden die invloed kunnen hebben op de bodemdieren. De data van benthos biomassa en dichtheid van de vooroever van de deelgebieden van Ameland en Schiermonnikoog laten zien dat er sprake is van grote jaar tot jaar variatie. Wanneer er ingezoomd wordt op de suppletie die heeft plaatsgevonden, wordt een lokale impact zichtbaar aan de buitenste rand van de suppletie en in een kleine zone van het direct ten oosten van de suppletie gelegen uitstralingsgebied. Na een periode van ongeveer twee jaar is er op deze locatie geen effect meer te onderscheiden in gemiddelde dichtheden en biomassa's van de bodemdieren ten opzichte van de overige locaties. Kijken we naar specifieke soorten, dan verschilt deze periode enigszins van soort tot soort. Zo laat het nonnetje pas na twee jaar gemiddelde dichtheden zien die vergelijkbaar zijn aan dichtheden voorafgaand aan de suppletie, terwijl dit bij de borstelworm Mage/ona johnstoni al na een jaar is. De zandzager Nepthys hombergii laat helemaal geen effect van de suppletie zien. Op basis van deze bevindingen is er geen aanleiding om de huidige ontwerprichtlijn voor de suppletiefrequentie van circa eens in de vier jaar aan te passen, omdat het herstel binnen deze periode plaatsvindt op Ameland. Doordat het een casestudie betreft, zijn resultaten niet op te schalen naar de gehele Nederlandse kust, hiertoe zal kustdekkend onderzoek moeten worden gedaan. Daarnaast zijn ook de cumulatieve effecten van het herhaaldelijk toepassen van zandsuppleties op de bodemdieren van de vooroever tot dusver onbekend. Ontwikkeling van het bodem/even in de vooroever na aan/eg van een onderwatersupp/etie - Case studie Ame/and en Schiermonnikoog


6 Deltares Titel Ontwikkeling van het bodemleven in de vooroever na aanleg van een onderwatersuppletie Opdrachtgever Rijkswaterstaat Project Kenmerk ZKS-0012 Pagina's 111 Versie Datum Auteur mrt 2016 S.A. Vergouwen "" MSc ir. H. Meijer Holzhauer Hoozemans Status definitief Ontwikkeling van het bodem/even in de vooroever na aan/eg van een onderwatersupp/etie - Case studie Ame/and en Schiermonnikoog
7
8 Inhoud Samenvatting Summary iii v 1 Inleiding 7 2 Methode Bemonstering Studiegebieden Laboratorium analyses Hypotheses Analyses Vergelijkingen Geanalyseerde parameters Statistiek Meteorologische parameters 20 3 Meteorologische omstandigheden Windsnelheden Temperatuur Neerslag 25 4 Sedimentsamenstelling 27 5 Resultaten macrobenthos Soortenaccumulatiecurves Ameland Midden Ameland Impact Ameland Uitstraling Schiermonnikoog Vergelijking tussen studiegebieden T0-vergelijking Gehele tijdsperiode Ontwikkeling per studiegebied Ameland Midden Ameland Impact Ameland Uitstraling Schiermonnikoog Losse soorten uitgelicht Macoma balthica Magelona johnstoni Nepthys hombergii Echinocardium cordatum 78 6 Conclusies 83 7 Doorvertaling naar de suppletiepraktijk 87 i
9 7.1 Suppletievolume Suppletiefrequentie Sedimentsamenstelling Kustdwarse positie Moment van suppleren 89 8 Discussie 91 9 Literatuur 95 Bijlage(n) A Jaarlijks verschil in D50 tov 2010 A-1 B Biomassa/dichtheid per soortgroep B-1 C Verspreiding Schiermonnikoog C-1 C.1 Biomassa per monsterlocatie C-1 C.2 Dichtheid per monsterlocatie C-4 C.3 Dichtheidverspreiding van losse soorten C-7 C.3.1 Macoma balthica C-7 C.3.2 Magelona johnstoni C-9 C.3.3 Nepthys hombergii C-12 C.3.4 Echinocardium cordatum C-14 ii
10 Samenvatting Om functies van de kust zoals veiligheid en recreatie te bewaren, wordt er sinds 1990 zand gesuppleerd langs de kust van Nederland, waarbij er gebruik gemaakt wordt van de natuurlijke processen langs de kust en zo het zandige karakter van de Nederlandse kust behouden wordt. Middels de zandsuppleties blijft de ligging van de kustlijn in 1990 (vertaald naar een zogenaamde BasisKustLijn (BKL)) gehandhaafd. In 2001 is, met het oog op duurzame bescherming, het beleid uitgebreid tot het compenseren van het zandvolume in het gehele kustfundament, i.e. van de binnenduinenrij tot de doorgaande -20 meter NAP-lijn. In totaal wordt jaarlijks 12 miljoen kubieke meter zand op de kust gesuppleerd. Dit gebeurd in een vierjarige cyclus, waarmee bijna de totale Nederlandse kust elke vier jaar wordt gesuppleerd. Hoewel zandsuppleties worden gezien als noodzakelijk voor het handhaven van de functies van de kust, bestaat er de zorg dat de huidige zandsuppleties van invloed zijn op het functioneren van het ecosysteem van de ondiepe kustzone. Om een antwoord te kunnen geven op de vraag of en hoe de veiligheid gehandhaafd kan worden door middel van zandsuppleties zonder dat het ecosysteem hier negatieve invloed van ondervindt, heeft Rijkswaterstaat met vier natuurbeschermingsorganisaties (Stichting de Noordzee, de Waddenvereniging, Stichting Duinbehoud en de Vogelbescherming) een samenwerkingsovereenkomst 1 opgesteld waarin is afgesproken om van 2009 t/m 2015 onderzoek te doen naar de mogelijke effecten van zandsuppleties op het kustecosysteem (Holzhauer et al. 2009). In dit kader is er in 2009 gekozen om de ontwikkeling van het bodemleven na een zandsuppletie te volgen voor een periode van vier jaar en daarmee een volledige suppletiecyclus te beslaan. Als studiegebied is Ameland-midden gekozen omdat voor dit gebied in een grote zandsuppletie gepland stond. De kust van Ameland wordt geregeld gesuppleerd, doordat de kustlijn over het algemeen een negatieve trend laat zien. Dit rapport geeft een overzicht van de ontwikkelingen van de bodemdieren van de vooroever op Ameland tussen Tijdens deze studie zijn er op Ameland twee deelgebieden onderzocht Ameland Impact, waar in 2011 is gesuppleerd, en Ameland Uitstraling, een gebied direct ten oosten van de suppletie, dat mogelijk indirecte effecten kan ondervinden van de suppletie. Schiermonnikoog is daarnaast door de tijd heen gevolgd als referentiegebied. Naast bodemdieren zijn de meteorologische omstandigheden en het sediment meegenomen in deze rapportage, om inzicht te geven in de fysische omstandigheden die invloed kunnen hebben op het bodemleven. De data van biomassa en dichtheid van het macrobenthos in de vooroever van Ameland en Schiermonnikoog, laten zien dat er sprake is van grote jaar tot jaar variatie. Tevens is gebleken dat het ongesuppleerde Schiermonnikoog een afwijkende bodemdiergemeenschap kent. Mogelijke redenen hiervoor zijn dat het kustprofiel van Schiermonnikoog veel vlakker is en een minder uitgesproken bankenpatroon heeft dan Ameland. Dit maakte het niet mogelijk om op basis van een directe vergelijking tussen Ameland en Schiermonnikoog een effect van de suppletie waar te nemen. 1 Convenant gesloten op 24 maart 2009 iii
11 Wanneer er ingezoomd wordt op de suppletiedieptezone wordt een lokale impact zichtbaar aan de buitenste rand van de suppletie en in een kleine zone van het direct ten oosten van de suppletie gelegen ongestoorde gebied. Na een periode van ongeveer twee jaar is er op deze locatie geen effect meer te onderscheiden ten opzichte van de overige monsterpunten. Kijken we naar specifieke soorten, dan verschilt deze periode enigszins van soort tot soort. Zo laat het nonnetje pas na twee jaar gemiddelde dichtheden zien die vergelijkbaar zijn aan dichtheden voorafgaand aan de suppletie, terwijl dit bij de borstelworm Magelona johnstoni al na een jaar is. De zandzager Nepthys hombergii laat helemaal geen effect van de suppletie zien. Op basis van deze bevindingen is er geen aanleiding om de huidige ontwerprichtlijnen voor de suppletiefrequentie, van circa eens in de vier jaar aan te passen. Opgemerkt wordt dat dit slechts een case studie is waarbij vooral gekeken is naar effecten die kunnen optreden na een suppletie. Om te toetsen of dezelfde uitkomsten gelden voor de gehele Nederlandse kust, zal ook hier studie naar gedaan moeten worden. De vergelijking met het toch zeer nabij gelegen Schiermonnikoog onderschrijft dit. Studie op de schaal van de gehele Nederlandse kust kan op verschillende manieren worden aangepakt, 1) in de vorm van het opvolgen van een suppletie op een nieuwe locatie, 2) door enkele vaste raaien langs de kust te kiezen waarlangs de bodemdierontwikkeling wordt opgevolgd ongeacht of er een suppletie wordt uitgevoerd of niet, of 3) door elke suppletie langs de Nederlandse kust kort op te volgen met enkele raaien aan monsters. Naast de opschaling naar de gehele Nederlandse kust zullen ook de cumulatieve effecten van jarenlang suppleren langs de kust op de samenstelling van de bodemdieren in ogenschouw genomen moeten worden. Lange termijn kustdekkend onderzoek zou hier meer inzicht in kunnen bieden. iv
12 Summary In order to maintain different functions of the Dutch coastal zone, such as safety against flooding and recreation, the coast is being nourished with sand since This dynamic coastal management makes use of natural processes along the shore and helps to maintain the sandy character of the Dutch coast. Sand nourishments allow the coastline that has been determined in 1990 (translated to a so called Basic Coast Line (BCL) to be preserved. Since 2001 Dutch policies regarding coastal protection have been extended to include the principle of sustainability: compensation of the sand volume of the entire coastal foundation, i.e. the inner dunes up until -20 meter below Amsterdam Ordnance Datum (Dutch: NAP). As a result, the coast is nourished with a total of 12 million cubic meters of sand each year. Nourishments are managed in a cycle ensuring that virtually each location on the Dutch coast is nourished once every four years. When possible, beach nourishments are avoided in favour of foreshore nourishments, based on the hypothesis that with foreshore nourishments sediment will distribute itself in the coastal zone by natural processes. Although sand nourishments are widely considered to be crucial for maintenance of the Dutch coast, there are concerns regarding the potential impact of sand nourishments on coastal ecosystem functioning. To assess the ecological impact of the current coastal nourishment regime and to investigate whether optimisations are possible to minimise negative effects, Rijkswaterstaat (Ministry of Infrastructure and Environment) is cooperating with four nature organisations (Stichting de Noordzee, Waddenvereniging, Stichting Duinbehoud and Vogelbescherming). In a mutual agreement 2 a decision was taken for a research project on ecological effects of coastal nourishments, for the time period (Holzhauer et al. 2009). Within this framework, the development of the benthic community has been monitored in relation to morphological changes following a sand nourishment. The project spanned a period of four years (starting in 2009), i.e. following an entire nourishment cycle. The island of Ameland has been chosen as a study area, because a large sand nourishment was planned here in The coastline of Ameland shows consistent erosion and landward retreat. It is therefore nourished on a regular basis. This report will provide an overview of the development of the benthic community of the foreshore of Ameland between This study has focussed on two areas of Ameland: Ameland Impact, which has been nourished in 2011, and Ameland Influence (Dutch: Ameland Uitstraling), an area directly east of the nourishment, where possible indirect effects of the nourishment could be expected. Together with these two study areas, the coast of a neighbouring island, Schiermonnikoog, has been monitored throughout time as a reference area. Benthos composition, meteorological conditions, bathymetry and sediment composition have been analysed, in order to provide insight in the physical circumstances that could affect benthic life. Data on biomass and density of benthos of the foreshore of the different study areas, show that there is large interannual variation. There were also marked differences in benthic communities between the Ameland sites prior to nourishment and the reference site of Schiermonnikoog despite the proximity of the islands and the apparent similarities between 2 Memorandum signed on the 24 th of March 2009 v
13 the locations. The foreshore profile of Schiermonnikoog is less steep and does not show such a pronounced shoal development, compared to Ameland. This may explain the differences between the sites. It is clear that a direct comparison between Schiermonnikoog and Ameland to assess the impact of the foreshore nourishment cannot be made. However, when we focus on the local depth range of the nourishment area we can demonstrate a local impact at the perimeter of the nourishment and in a small area due east of the nourishment location. However, an impact of the nourishment is visible in both biomass and density very locally at the outer border of the sand nourishment in Ameland Impact, and in a small part of the neighbouring area (Ameland Influence) directly east of the nourishment. After roughly two years, affected locations are indistinguishable from the other sample points. There is a difference in the observed impact of the nourishment between species. For bivalve Macoma balthica it takes two years for densities to recover to levels similar to the period prior ot the nourishment. For the shovelhead worm, Magelona johnstoni, this happens already after one year. The marine catworm Nephtys hombergii does not show an impact of the nourishment at all. Based on these findings, there is no clear argument to alter the current design directive for a nourishment frequency of once every four years. It should be noted that this is merely a case study, focussing on the direct, local effects. Testing whether these results are valid for the entire Dutch coast, requires research on a larger geographical scale. There are different ways to approach this: 1) monitoring the effects of several nourishments at different locations, 2) monitoring the distribution of benthic species at fixed locations along the Dutch coast, independent of nourishments., 3) monitoring the impact of each nourishment along the Dutch coast with a fixed number of samples and moments in time. Together with the spatial upscaling of research to the entire Dutch coast, the cumulative effects of repeated sand nourishments over time on benthic species should be taken into account. It is not really possible to get a handle on these effects by studying the few locations along the Dutch coast that are not regularly nourished (such as Schiermonnikoog), as these sites differ in several aspects from the sites that are nourished. Long-term research that covers the entire coast should provide more insight in these potential effects. vi

14 1 Inleiding Sinds 1990 worden er langs de Nederlandse kust zandsuppleties uitgevoerd met als doel het bewaren van functies van de kust zoals veiligheid en recreatie. In 2001 is, met het oog op duurzame bescherming, het beleid uitgebreid tot het compenseren van het zandvolume in het gehele kustfundament, i.e. van de binnenduinenrij tot de doorgaande -20 meter NAP-lijn. Het benodigde zand wordt op het strand of in een smalle zone (-5 tot -8 m NAP) onderwater aangebracht. Jaarlijks wordt er langs de hele Nederlandse kust ongeveer 12 Mm 3 zand aangebracht (zie figuur 1.1 voor de suppletiehoeveelheden per jaar sinds 1991) Het benodigde zand wordt gewonnen buiten de doorgetrokken -20 meter diepte lijn. De natuurlijke zandige kust wordt zo dynamisch gehandhaafd en is op deze manier veilig en kan ruimte bieden aan diverse gebruiksfuncties. Figuur 1.1 Uitgevoerde suppleties voor regulier onderhoud van de kustlijn en aantal overschrijdingen BKL sinds *Voor 2015 is uitgegaan van de uitgevoerde kuubs tot 1 november 2015, aangevuld met de verwachte realisatie t/m december (Ministerie van Infrastructuur en Milieu 2015). Hoewel zandsuppleties worden gezien als noodzakelijk voor het handhaven van de functies van de kust, bestaat er de zorg dat de huidige zandsuppleties van invloed zijn op het functioneren van het ecosysteem van de ondiepe kustzone. Op de vooroevers van de Nederlandse kust komen o.a. macrobenthische gemeenschappen voor bestaande uit verschillende schelpdieren, wormen en kreeftachtigen. Deze macrofauna vormt een belangrijke schakel in de voedselketen, vooral als prooi voor vogels en (juveniele) vissen. De aanwezigheid van deze organismen wordt in grote mate bepaald door de geschiktheid van het gebied om zich er te vestigen en te overleven. Of een gebied geschikt is wordt o.a. bepaald door de aanwezige getijdestroming, golven, bodemvorm, sedimentsamenstelling en diepte. Door een zandsuppletie worden de abiotische en biotische factoren en processen ter plekke en in de nabije omgeving tijdelijk of voor een langere tijd beïnvloed (Speybroeck et al. 2006). Om een antwoord te kunnen geven op de vraag of en hoe de veiligheid gehandhaafd kan worden door middel van zandsuppleties zonder dat het ecosysteem hier negatieve invloed van ondervindt, heeft Rijkswaterstaat met vier natuurbeschermingsorganisaties (Stichting de Noordzee, de Waddenvereniging, Stichting Duinbehoud en de Vogelbescherming) een 7
15 samenwerkingsovereenkomst 3 opgesteld waarin is afgesproken om van 2009 t/m 2015 onderzoek te doen naar de mogelijke effecten van zandsuppleties op het kustecosysteem (Holzhauer et al. 2009). In dit kader is er in 2009 gekozen om binnen het programma Kennis voor Primaire Processen Beheer & Onderhoud Kust de ontwikkeling van het bodemleven na een zandsuppletie te volgen voor een periode van vier jaar en daarmee een volledige suppletiecyclus te beslaan. Als studiegebied is Ameland-midden gekozen omdat voor dit gebied in 2010/2011 een grote zandsuppletie gepland stond. (zie figuur 1.2 voor de hoeveelheden zandsuppletie lang de gehele kust van Nederland per kustvak per jaar). Figuur 1.2 Totale hoeveelheid uitgevoerde zandsuppleties voor onderhoud van de kustlijn van de periode 2005 t/m 2015, per kustvak. Voor 2015 is uitgegaan van de uitgevoerde kuubs tot 1 november 2015, aangevuld met de verwachte realisatie t/m december 2015 (Ministerie van Infrastructuur en Milieu 2015). Niet alleen op Ameland-midden worden er zandsuppleties aangebracht. De gehele kust van Ameland wordt geregeld gesuppleerd, doordat de kustlijn over het algemeen een negatieve trend laat zien. (zie figuur 1.3 Ameland voor de kustlijn van 2015). In het westelijke deel wordt de kustlijn van Ameland beïnvloed door het landwaarts verplaatsen van de geul, het Borndiep, en de zandplaat, het vroegere Bornrif, dat zich langzaam van het westen naar het oosten van het eiland verplaatst en nu een geheel vormt met het strand. Ook hier zijn verschillende zandsuppleties uitgevoerd. Voorbij Ballum verplaatst de kustlijn zich zeewaarts als gevolg van de oostwaartse beweging van de zandplaat het Bornrif. In het midden en oosten van Ameland heeft de afgelopen jaren een kustachteruitgang plaatsgevonden. Al in 1998 is hier de eerste vooroeversuppletie uitgevoerd waarbij 1,5 miljoen m 3 zand werd aangebracht. Sindsdien zijn er in 2003, 2006, en 2015 vooroeversuppleties al dan niet in combinatie met strandsuppleties aangebracht. In het najaar van 2011 zijn hier twee 3 Convenant gesloten op 24 maart



16 zandsuppleties afgerond. Een vooroeversuppletie, welke is opgevolgd in deze studie, en een strandsuppletie. In de huidige toetsing van 2015 is de BKL geheel oostelijk op het eiland opnieuw overschreden en voor het midden van Ameland is de trend opnieuw landwaarts gericht. In 2015 is hier wederom een strandsuppletie gecombineerd met een vooroeversuppletie uitgevoerd. De effecten van de strandsuppleties worden in dit rapport verder niet beschouwd. Als referentiegebied voor het studiegebied Ameland-midden is Schiermonnikoog gekozen. Er is op Schiermonnikoog nog nooit gesuppleerd. De kustlijn van Schiermonnikoog ligt minimaal 120 meter zeewaarts van de BKL. Ondanks dat er een achteruitgang wordt waargenomen zijn er tot en met 2020 geen overschrijdingen van de BKL te verwachten. Ameland Schiermonnikoog Figuur 1.3 Kustlijn van Ameland en Schiermonnikoog in 2015 (Ministerie van Infrastructuur en Milieu 2015) 9
17 In het kader van deze studie is in 2009 (Goudswaard et al. 2009, Wijsman et al. 2010) en 2010 (Van de Moortel et al. 2010, Vanagt et al. 2010, Heusinkveld et al. 2011, Vanagt et al. 2011) een T0-bemonstering van het bodemleven van de vooroever uitgevoerd om de beginsituatie voorafgaand aan de suppletie in 2010/2011 te bepalen. Vervolgens is er na de aanleg van de onderwatersuppletie vier keer in het najaar het bodemleven in de vooroever bemonsterd in 2011 (Verduin et al. 2011, Leewis et al. 2012, Verduin et al. 2012), 2012 (Holzhauer et al. 2012, Faasse et al. 2013, Heusinkveld 2013), 2013 (Heusinkveld 2014) en 2014 (Van Dalfsen et al. 2014a, Van Dalfsen et al. 2014b, Verduin et al. 2014b, Vergouwen et al. 2015). In 2013 is een eerste interim rapportage opgesteld over de waargenomen ontwikkelingen van 2009 tot en met 2012 (Holzhauer et al. 2014). De ontwikkeling van de bodemdiergemeenschap van de vooroever over de jaren is nog niet eerder geanalyseerd. Het doel van deze rapportage is om een overzicht te geven van de ontwikkelingen van het macrobenthos voor Ameland en Schiermonnikoog van jaar tot jaar in relatie tot de onderwatersuppletie aangelegd op Ameland midden in 2010/
18 2 Methode 2.1 Bemonstering Tussen 2009 en 2014 hebben er jaarlijkse bemonsteringen plaatsgevonden in het najaar op de vooroever van Ameland en Schiermonnikoog van de bodemdiersamenstelling, bodemhoogte en sedimentsamenstelling in en nabij het gesuppleerde gebied. Een bemonstering in het najaar geeft een geconsolideerd beeld van de gevestigde individuen. De biomassa is hoog omdat de organismen in de productie van de zomer hebben kunnen benutten, er nog geen lage temperaturen zijn opgetreden en de eierproductie nog niet is geweest. Je kunt grofweg stellen dat ze zich klaar hebben gemaakt voor de winter. Het najaarsbeeld is goed voor het monitoren van trendmatige veranderingen in diversiteit, aantallen en biomassa. In het voorjaar zijn deze parameters op de termijn van weken tot maanden een stuk variabeler. Stratum Code Omschrijving 1 H1 De kustwaarts gelegen helling t.o.v. van de eerste trog 2 Tr1 De bodem van de eerste trog 3 B1 De top van de eerste brekerbank 4 H2 De kustwaarts gelegen helling t.o.v. van de buitenste trog 5 Tr2 De bodem van de buitenste trog 6 B2 De top van de buitenste brekerbank 7 H3 De zeewaarts gelegen helling t.o.v. van de top van de buitenste brekerbank, binnen suppletiegebied 8 H4 De buitenste helling, op circa 7 m diepte, net binnen het suppletiegebied 9 H5 De buitenste helling, op circa 9 m diepte 10 HD De buitenste helling, op circa 10 m diepte Figuur 2.1 Schematische dwarsdoorsnede van de vooroever met de monsterlocaties en de morfologische beschrijving van de locatie. Het rode vak geeft de monsterlocaties in het suppletiegebied weer. 11
19 De bemonsteringen zijn uitgevoerd met behulp van een Van Veenhapper vanaf een ondiepstekend schip over kustdwarse raaien met een onderlinge afstand van circa 500 m. Op elke raai zijn circa 10 monsters genomen op een vooraf bepaalde locatie, aan de hand van de dan geldende morfologie (Figuur 2.1). De reden om de monsterpunten geheel volgens de geldende morfologie in te plannen komt voort uit de veronderstelling dat de verspreiding van de bodemdieren voor een belangrijk deel wordt bepaald door de aanwezige morfologische kenmerken van de ondiepe vooroever, zoals troggen en banken. Het meest diepe monsterpunt ligt op circa 3 km uit de kust gerekend vanaf de laagwaterlijn. 2.2 Studiegebieden De studiegebieden, suppletielocatie en het aantal monsters per bemonstering zijn weergegeven in figuur 2.2. en tabel 2.1. Op Ameland zijn er drie studiegebieden en op Schiermonnikoog één. Ameland Midden (AM: RSP ) Ameland Midden is in 2009 en 2014 bemonsterd. Er heeft een vooroeversuppletie plaatsgevonden in mei t/m augustus Ameland Midden is enkel het jaar voorafgaand aan de suppletie bemonsterd en vier jaar later. In 2015 is er opnieuw een vooroeversuppletie uitgevoerd (Figuur 2.2-A). Ameland Impact (AI: RSP 17-20) Het gebied Ameland Impact is bemonsterd in 2009, 2010, 2011, 2012, 2013 en Ameland Impact is gesuppleerd in januari t/m juli 2011 (Figuur 2.2-A). Ameland Uitstraling (AU: RSP 20-23) Het gebied Ameland Uitstraling is nooit gesuppleerd maar ligt direct naast het suppletiegebied. De verwachting is dat hier een uitstralende werking is van de naast gelegen suppletie. Ameland Uitstraling is bemonsterd in 2009, 2010, 2011, 2012 en 2013 (Figuur 2.2- A). Schiermonnikoog (SR: RSP 11-14) De vooroever van Schiermonnikoog is nooit gesuppleerd en wordt ook niet direct beïnvloed door de uitgevoerde suppletie. Schiermonnikoog wordt gebruikt als referentiegebied. Op basis van de analyse van de monsters in 2010 is bepaald dat de bodemdiersamenstelling van Ameland en Schiermonnikoog voldoende overeen kwamen om Schiermonnikoog als referentiegebied te kiezen (Vanagt et al. 2011). Uit de latere analyses blijkt echter dat Schiermonnikoog vrijwel altijd afwijkt van Ameland. Als reden wordt hiervoor gegeven dat de vooroever van Schiermonnikoog ondieper is en een flauwer profiel heeft dan Ameland; respectievelijk circa -8 m NAP vs. circa -10 m NAP op 2500 m uit de kust. Daarnaast is het dubbele bank-trog systeem meer geprononceerd in de vooroever van Ameland dan bij Schiermonnikoog (Holzhauer et al. 2014). Zie voor meer details de afzonderlijke jaarrapportages en de resultaten beschreven in deze rapportage. De bemonstering van Schiermonnikoog heeft plaatsgevonden in 2010 t/m 2014 (Figuur 2.2-B). 4 RSP = Rijksstrandpaal. Dit is de nummering van de kust van Rijkswaterstaat en komt overeen met de palen op het strand en de raaien zoals weergegeven in bijvoorbeeld het kustlijnkaartenboek. 12
 Ameland: AM = Ameland Midden, AI = Ameland Impact, AU = Ameland Uitstraling.")

20 AM AI AU A) SR Figuur 2.2 Overzicht onderzoekslocaties. Figuur A) Ameland: AM = Ameland Midden, AI = Ameland Impact, AU = Ameland Uitstraling. Figuur B) Schiermonnikoog: SR = Schiermonnikoog Referentie. Monsterlocaties: grijs = bemonstering 2009, rood = bemonstering 2010, 2011, 2012 en 2013, groen = bemonstering Suppletie: oranje vak = suppletielocatie mei t/m oktober ,9 milj. m 3, gele vak = suppletielocatie mei t/m augustus ,1 milj. m 3 en rode vak = suppletielocatie januari t/m juli ,6 milj. m 3. B) 13
21 Tabel 2.1 Overzicht momenten van bemonsteren per studiegebied inclusief het aantal monsters (mb = macrobenthos, Sed = sediment) en momenten van suppleren (grijs gemarkeerd). *Ameland Overig is geen studiegebied. Hier is wel gesuppleerd en alleen in 2009 zijn hier monsters genomen t.b.v. van de vergunning om uit te sluiten dat er schelpdierbanken aanwezig waren. monsteren/ suppleren Ameland Overig* RSP Ameland Midden RSP Studiegebieden Ameland Impact RSP Ameland Uitstraling RSP Schiermonnikoog RSP T t/m 5 aug 2009 (grid) 33 mb 5 sed 14 mb 5 sed 17 mb 6 sed 6 mb 13 sed Mei t/m okt 2010 RSP 11,4 t/m 14 (1,9 Milj m 3 ) Mei t/m aug 2010 RSP 14 t/m 17 (1,1 Milj m 3 ) T aug t/m 3 sept 2010 (6 * 3 raai) 60 mb 19 sed 60 mb 20 sed 68 mb 13 sed Jan t/m juli 2011 RSP 17 t/m 20 (1,6 Milj m 3 ) T1 24 t/m 30 sept 2011 (6 * 3 raai) 60 mb 27 sed 56 mb 29 sed 60 mb 30 sed T2 1 okt t/m 3 okt 2012 (6 * 3 raai) 66 mb 33 sed 61 mb 31 sed 17 mb 7 sed T3 3 t/m 6 sept 2013 (6 * 3 raai) 66 mb 22 sed 66 mb 22 sed 60 mb 21 sed T t/m 5 aug 2014 (3*3 raai) 36 mb 10 sed 36 mb 10 sed 36 mb 10 sed Jan t/m aug 2015 RSP 11,4 t/m 16 (2,0 Milj m 3 ) 2.3 Laboratorium analyses Nadat de bodemdiermonsters en sedimentmonster zijn verzameld zijn ze overgebracht naar het laboratorium. Bodemdieren De bodemdiermonsters zijn eerst uitgezocht waarna alle organismen zoveel mogelijk tot op soortsniveau zijn gedetermineerd en tot slot is de biomassa bepaald. Voor het uitzoeken zijn de monsters eerst gekleurd met Bengaals roze waarna de monsters grondig zijn gespoeld om zoveel mogelijk van de fixatievloeistof te verwijderen, waarmee de monsters in het veld waren gefixeerd. Vervolgens zijn de monsters uitgezocht en zijn de organismen samengevoegd naar hogere taxonomisch niveaus. Het determineren van de organismen is gebeurd met behulp van een binoculaire microscoop en de meest recente determinatieliteratuur. Voor nomenclatuur is de TWN-lijst en indien deze geen uitsluitsel geeft, het World Register of Marine Species (WoRMS) ( gevolgd. De biomassa, in gram asvrij drooggewicht (g ADG), is via conversie bepaald. Dit is overeenstemming met de methode gehanteerd voor de MWTL benthosbemonsteringen van Rijkswaterstaat (Templeman et al. 2009). Opgemerkt wordt dat de laboratorium analyses van de verschillende jaren zijn door verschillende partijen geanalyseerd. In 2009 door IMARES, in 2010 (Van de Moortel et al. 2010), 2012 (Faasse et al. 2013) en 2013 (Faasse et al. 2014) door ecoast en in 2011 en 2014 (Verduin et al. 2014a) door Grontmij. 14



22 Sediment De sedimentmonsters zijn donker en koel bewaard en bij aankomst in het laboratorium ingevroren op -16 C tot analyse van de korreldiameter met behulp van de Malvern en het bepaling van het organisch stof gehalte met behulp van een TGA. Ook hier zijn gedurende de jaren de monsters door verschillende partijen geanalyseerd. In 2010, 2011 (Leewis 2011) door de Vrije Universiteit van Amsterdam, in 2012 en 2013 door de Universiteit van Gent en in 2014 (Ibanez 2014, Ibanez et al. 2014) door Deltares. 2.4 Hypotheses Het doel van deze rapportage is om een overzicht te geven van de ontwikkelingen van het macrobenthos voor Ameland en Schiermonnikoog van jaar tot jaar in relatie tot de onderwatersuppletie aangelegd in op Ameland midden in 2010/2011. Om antwoord te kunnen geven op deze onderzoeksvraag zijn drie hypotheses geformuleerd. Allereerst wordt er gekeken of het macrobenthos van de studiegebieden van Ameland en Schiermonnikoog van elkaar verschillen op grote schaal, door naar het gemiddelde beeld van alle jaren te kijken. De gebieden hebben allen een andere historie met betrekking tot zandsuppleties. Deze analyse biedt inzicht of de bodemdiergemeenschap van de verschillende studiegebieden met afwijkende historie op elkaar lijken op grote schaal, over meerdere jaren. H0-1: Alle studiegebieden zijn gelijk in dichtheid, biomassa en diversiteit onderling over de hele periode Vervolgens wordt er per studiegebied (elk met een eigen historie) gekeken of er over de jaren een ontwikkeling is geweest, door te analyseren of binnen een studiegebied het macrobenthos van verschillende jaren van elkaar afwijken. Doordat alle gebieden een andere historie hebben met betrekking tot suppleties, kan dit voor verschillen in de bodemdiersamenstelling hebben gezorgd van jaar tot jaar. Hiertoe is de volgende hypothese geformuleerd: H0-2: De afzonderlijke studiegebieden komen overeen tussen de jaren in dichtheid, biomassa en diversiteit. 15
23 Tot slot is ook specifiek naar de suppletiezone gekeken van Ameland Impact in vergelijking tot het niet gesuppleerde Ameland Uitstraling en Schiermonnikoog. Hoewel niet in elk gebied gesuppleerd is, is er wel in de suppletiezone bemonsterd. De ontwikkeling van deze zone over de jaren in verschillende studiegebieden (gesuppleerd en niet gesuppleerd) kan de directe effecten en lange termijneffecten meer inzichtelijk maken. Hiertoe wordt ook een aantal soorten dat uit eerdere rapportages een effect bleek te ondervinden, of waarbij een effect van de suppletie verwacht wordt, geanalyseerd. Deze soorten zijn het nonnetje Macoma balthica (een langlevende soort die mogelijk benadeeld wordt door suppleties), Nephtys hombergii (de zandzager; een opportunistische worm), Magelona johnstoni (opportunistische borstelworm) en Echinocardium cordatum (zeeklit, een grote soort die zich makkelijk verplaatst). H0-3: Er is geen verschil in de ontwikkeling van het macrobenthos in de suppletiezone in het gesuppleerde Ameland Impact, ten opzichte van het niet-gesuppleerde Ameland Uitstraling en Schiermonnikoog. Hieronder wordt een overzicht weergegeven van de deelvragen per hypothese en de analyse die is uitgevoerd om de verschillende deelvragen te beantwoorden. Tabel 2.2 Deelvragen en analyses bij hypotheses Hypothese Deelvraag Analyse H0-1: De studiegebieden verschillen niet onderling over de hele periode Verschillen studiegebieden onderling in biomassa (g ADG/m 2 ) op een grotere tijdschaal (alle bemonsterde jaren)? Verschillen studiegebieden onderling in dichtheid (n/m 2 ) op een grotere tijdschaal (alle bemonsterde jaren)? Verschillen studiegebieden onderling in diversiteit op een grotere tijdschaal (alle bemonsterde jaren)? Gemiddelde biomassa (g ADG/m2) van alle studiegebieden van de bemonsterde jaren in een staafdiagram, visuele inspectie. One-way ANOVA (afhankelijk: gemiddelde dichtheid per monster (n/m2); onafhankelijk: studiegebied). Levene's test voor homogeniteit. Post-Hoc Tukey Kramer paarsgewijze test. Gemiddelde dichtheid (n/m 2 ) van alle studiegebieden van de bemonsterde jaren in een staafdiagram, visuele inspectie. One-way ANOVA (afhankelijk: gemiddelde biomassa per monster (g ADG/m 2 ); onafhankelijk: gebied). Levene's test voor homogeniteit. Post-Hoc Tukey Kramer paarsgewijze test. Shannon-Wiener s en Pielou's index voor diversiteit per studiegebied op basis van dichtheid (n/m 2 ). Gemiddeld aantal soorten per monster (n/m 2 ). One-way ANOVA (afhankelijk: gemiddelde Shannon-Wiener diversiteit per monster; onafhankelijk: gebied). Levene's test voor homogeniteit. Post-Hoc Tukey Kramer paarsgewijze test. 16
24 H0-2: De studiegebieden verschillen niet tussen de jaren. H0-3: De studiegebieden verschillen niet in de suppletiezone tussen de jaren Verschillen afzonderlijke studiegebieden tussen de jaren in biomassa (g ADG/m 2 )? Verschillen studiegebieden tussen de jaren in dichtheid (n/m 2 )? Verschillen studiegebieden tussen de jaren in diversiteit? Verschillen afzonderlijke studiegebieden tussen de jaren in biomassa (g ADG/m 2 ) in het suppletiegebied Verschillen studiegebieden tussen de jaren in dichtheid (n/m 2 ) in de suppletiezone? Verschillen studiegebieden tussen de jaren in diversiteit? Gemiddelde biomassa (g ADG/m 2 ) per jaar van de bemonsterde jaren in een staafdiagram voor elk studiegebied, visuele inspectie. One-way ANOVA (afhankelijk: gemiddelde biomassa per monster (g ADG/m 2 ); onafhankelijk: jaar). Levene's test voor homogeniteit. Post-Hoc Tukey Kramer paarsgewijze test. Gemiddelde dichtheid (n/m 2 ) per jaar in een staafdiagram voor elk studiegebied, visuele inspectie. One-way ANOVA (afhankelijk: gemiddelde dichtheid per monster (n/m 2 ); onafhankelijk: jaar). Levene's test voor homogeniteit. Post-Hoc Tukey Kramer paarsgewijze test. Shannon-Wiener s en Pielou's index voor diversiteit per jaar op basis van dichtheid (n/m 2 ). Gemiddeld aantal soorten per monster (n/m 2 ) per jaar, voor elk studiegebied One-way ANOVA (afhankelijk: gemiddelde Shannon-Wiener diversiteit per monster; onafhankelijk: jaar). Levene's test voor homogeniteit. Post-Hoc Tukey Kramer paarsgewijze test. Gemiddelde biomassa (g ADG/m 2 ) per jaar in het suppletiegebied voor elk studiegebied, aan de hand van het aantal maanden na de suppletie op AI Gemiddelde dichtheid (n/m 2 ) per jaar in het suppletiegebied voor elk studiegebied, aan de hand van het aantal maanden na de suppletie op AI Two-factor crossed ANOSIM in Primer V6 en een MDS plot Shannon-Wiener s en Pielou's index voor diversiteit per jaar op basis van dichtheid (n/m 2 ) in het suppletiegebied. 2.5 Analyses Vergelijkingen Vergelijking tussen gebieden Vergelijkingen tussen verschillende studiegebieden zijn gemaakt op basis van gemiddelde (dichtheid en biomassa) per monster, waarbij de monsters van alle jaren zijn meegenomen. Niet alle gebieden zijn een gelijk aantal jaren bemonsterd en binnen de jaren verschilt ook het monsteraantal. Hiervoor is gecorrigeerd in de analyse. Tevens is er een T0 vergelijking van de gebieden gemaakt in In de T0 van 2010 is Ameland Midden niet meegenomen, omdat dit gebied al in 2010 is gesuppleerd. Vergelijking binnen gebieden Vergelijkingen tussen jaren binnen studiegebieden, zijn gedaan op basis van een gemiddelde (biomassa, dichtheid) per monster per jaar. In 2009 is de biomassa niet bepaald (Tabel 2.3). 17
25 Biomassa Dichtheid Biomassa Dichtheid Tabel 2.3 Parameter Overzicht van monsteraantal per studiegebied per jaar, voor de parameters biomassa en dichtheid. Studiegebied Jaar AM AI AU SR Totaal Vergelijkingen tussen suppletiedieptezones Hoewel Ameland Uitstraling en Schiermonnikoog niet gesuppleerd zijn, wordt er toch specifiek naar de zone gekeken die gesuppleerd zou worden als er een vooroeversuppletie zou worden uitgevoerd. Deze zone is het buitenste deel van de brekerbankzone en is direct te vergelijken met de suppletiedieptezone van Ameland Impact, waar wel gesuppleerd is. De zone ligt over het algemeen tussen de -5 en -8 m diep, en wordt in dit rapport suppletiedieptezone genoemd (ongeacht of er gesuppleerd is of niet). De suppletiedieptezone van Ameland Impact, Ameland Uitstraling en Schiermonnikoog wordt geanalyseerd op basis van dichtheid en biomassa in aantal maanden sinds de suppletie op Ameland Impact. De suppletiedieptezone zijn de volgende strata: Helling 2 (stratum 4), Trog 2 (stratum 5), Bank 2 (stratum 6) en Helling 3 (stratum 7) (zie figuur 2.1). In 2011 zijn de strata van Ameland Uitstraling en Ameland Impact anders gelabeld (in de dataset) en ligt de suppletiedieptezone in Suppletiegebied 1, Bank 2, Helling 1 en Helling 3. De geografische ligging van deze zone verschilt niet in Het monsteraantal in de suppletiedieptezone van de verschillende studiegebieden is niet in alle jaren geheel gelijk. In 2009 zijn er te weinig monsters genomen in de suppletiedieptezone, daarom wordt dit jaar buiten beschouwing gelaten. Tevens wordt voor Schiermonnikoog het jaar 2012 buiten beschouwing. Wegens zeer slechte weersomstandigheden zijn hier slechts een aantal monsters genomen kunnen worden (n=5). Tabel 2.4 Parameter Overzicht van monsteraantal per studiegebied enkel in de suppletiedieptezone van Ameland Impact en Ameland Midden, en de (niet-gesuppleerde) overeenkomstige raaien van Ameland Uitstraling en Schiermonnikoog per jaar, voor de parameters biomassa en dichtheid. Studiegebied suppletiedieptezone Jaar AM AI AU SR Totaal
26 Losse soorten uitgelicht Doordat er voor bepaalde soorten met name een effect wordt verwacht van de suppletie, zijn er een aantal soorten uitgelicht waar nader op wordt ingezoomd. Deze soorten zijn: Macoma balthica (nonnetje): belangrijke vertegenwoordiger van de langlevende soorten Nephtys hombergii (zandzager): opportunistische aas-etende soort die mogelijk op korte termijn een voordeelpositie kan genieten. Echinocardium cordatum (zeeklit): verplaatst zich makkelijk en heeft een grote invloed op de totale biomassa. Magelona johnstoni (borstelworm): opportunistische soort die mogelijk op korte termijn een voordeelpositie kan genieten vanwege de snelle rekrutering. Van een deze individuele soorten wordt zowel de dichtheid, biomassa als de biomassa/dichtheid (een maat voor de biomassa per individu) weergegeven. Hierbij is een onderscheid gemaakt tussen voorkomen in de suppletiedieptezone, de ondiepe zone en de zeewaartse zone Geanalyseerde parameters Soortenaccumulatiecurve Voorafgaande aan de analyses is gekeken of het aantal genomen monsters representatief is voor de bodemdiergemeenschap van de studiegebieden. Een soortenaccumulatiecurve geeft weer of het aantal genomen monsters representatief is voor de aanwezige soorten in een gebied. Voor elk gebied is een soortenaccumulatiecurve gemaakt in Primer v6. Bij een asymptotische soortenaccumulatiecurve zijn er voldoende monsters genomen om het gebied te representeren en zullen extra monsters niet veel nieuwe soorten bevatten. Biomassa De gemiddelde biomassa (g ADG/m 2 ) van monsters per soort wordt weergegeven in een staafdiagram per gebied (H0-1) en per jaar (H0-2) voor de verschillende studiegebieden, aan de hand van de meest dominante soorten en een groepering van de overige soorten. Vanwege de verschillen in monsters en bemonsterde jaren, wordt er per soort een gemiddelde waarde berekend op basis van de genomen monsters (n). Voor biomassa worden alle analyses met en zonder Amerikaanse zwaardschede (Ensis directus) en zeeklit (Echinocardium cordatum) gedaan, vanwege hun relatief hoge biomassa per individu. In 2009 is de biomassa niet bepaald en wordt deze dus ook niet meegenomen. Dichtheid De gemiddelde dichtheid (n/m 2 ) van monsters per soort wordt weergegeven per gebied (H0-1) en per jaar (H0-2) voor de verschillende studiegebieden, in een staafdiagram aan de hand van de meest dominante soorten en een groepering van de overige soorten. Vanwege de verschillen in monsters en bemonsterde jaren, wordt er per soort een gemiddelde waarde berekend op basis van de genomen monsters (n). Voor dichtheid zijn ook verspreidingsfiguren gemaakt waarbij de gemiddelde dichtheid per monsterpunt weergegeven is op de coördinaten van dit monster. Dit is vooral relevant om de effecten van de suppletie inzichtelijk te maken. Diversiteit en species richness De species richness geeft weer hoeveel unieke soorten er zijn gevonden in een gebied. De diversiteit van verschillende gebieden wordt weergegeven aan de hand van de Shannon- 19
27 Wiener diversiteitsindex (berekend in Primer6). Deze wordt bepaald op basis van dichtheid per soort (n/m 2 ) per monster met behulp van de volgende formule: Waarbij H de Shannon Wiener diversiteitsindex is en p i de proportie van individuen per soort. De Shannon-Wiener diversiteitsindex is tevens als maat voor diversiteit gehanteerd in voorgaande rapportages over macrobenthos van Ameland (Goudswaard et al. 2009, Vanagt et al. 2011, Verduin et al. 2012, Holzhauer et al. 2014, Vergouwen et al. 2015). Verschil in diversiteit is statistisch getest op basis van de Shannon-Wiener diversiteitsindex per monster per gebied (H0-1) of per jaar binnen gebied (H0-2), met behulp van een one-way Analysis of Variance (ANOVA). Tevens is de Pielou evenness bepaald. De Pielou evenness is een index met een range van 0 tot 1, en zegt iets over hoe de verdeling in aantallen van de voorkomende soorten in het gebied is. Met andere woorden: of de soorten allemaal in min of meer gelijke aantallen voorkomen, of sterk verschillen in aantal. Hoe lager de variatie in het voorkomen van soorten, hoe dichter de waarde bij 1 ligt. De Pielou evenness index is bepaald in Primer-6, met behulp van de volgende formule: Waarbij J de Pielou evenness index is, H de Shannon Wiener diversiteitsindex en S het totaal aantal soorten in een monster Statistiek Verschillen tussen gebieden (H0-1) en jaren in verschillende studiegebieden (H0-2) zijn geanalyseerd in IBM SPSS Statistics 22 met behulp van een Analysis of Variance (ANOVA). Waarden zijn vooraf getransformeerd met een log10 functie. Doordat de monsteraantallen verschillen tussen gebieden en de data niet voldoen aan de voorwaarden voor een meer standaard Tukey Kramer post-hoc test, is een Games-Howell post-hoc paarsgewijze test uitgevoerd. Het betrouwbaarheidsinterval is op 95% gesteld. Verschillen tussen de suppletiedieptezone van de studiegebieden in verschillende jaren worden geanalyseerd met behulp van een Analysis of Similarity (ANOSIM) in Primer6, waarbij een two-way crossed design is gebruikt met als factoren studiegebied en aantal maanden sinds suppletie op AI. ANOSIMS worden uitgevoerd op basis van gemiddelde per unieke soort per monsters in de suppletiedieptezone Meteorologische parameters Meteorologische omstandigheden zijn gemeten ter hoogte van Terschelling voor de jaren 2009 tot en met Weergegeven parameters zijn neerslag, windsnelheden en temperatuur van de lucht en het water. Gegevens zijn afkomstig van waterbase en het KNMI. 20

 storm (maar wel veel lokaler).")
28 3 Meteorologische omstandigheden In dit hoofdstuk worden de meteorologische parameters gemeten bij Terschelling weergegeven ter illustratie van de grootschalige fysische omstandigheden in de periode van bemonstering. Het macrobenthos kan beïnvloed worden door uitzonderingen in de meteorologische omstandigheden. Zo kan een storm ervoor zorgen dat de brekerbanken omgewoeld worden en verplaatsen. Individuen raken los, of begraven en turbiditeit is tijdelijk verhoogd (Birchenough et al. 2011). Er wordt vaak gesuggereerd dat het effect van een suppletie vergelijkbaar is met het effect van een (grote) storm (maar wel veel lokaler). Windsnelheden zijn weergegeven om inzicht te krijgen in de mate waarin dergelijke verstorende events hebben plaatsgevonden. Ook extreme temperaturen kunnen de levenscyclus van soorten beïnvloeden (Hiscock et al. 2004), waardoor deze omstandigheden zijn meegenomen. Tot slot zou een grote hoeveelheid neerslag en input van zoetwater mogelijk tot zoutstress kunnen leiden bij macrobenthische soorten (Telesh et al. 2013). Om deze redenen zijn de meteorologische parameters windsnelheden, temperatuur en neerslag weergegeven. 3.1 Windsnelheden In figuur 3.1 zijn de etmaal-gemiddelde windsnelheden voor de jaren 2009 t/m 2014 weergegeven. Dit betekent dat er gedurende de dag langdurig hogere lagere windsnelheden zijn opgetreden. De hoogste uurgemiddelde snelheid is daarom ook weergegeven in de grafieken. 21




29 Figuur 3.1 Windsnelheden ter hoogte van Terschelling voor de jaren 2009 tot en met 2014.De blauwe lijn geeft de monsterperiode aan. Rode datapunten geven het maximaal uurgemiddelde van de windsnelheid weer. Gele ovalen geven stormevents voor de suppletie weer. 22
30 Tabel 3.1 Type wind met bijbehorende windsnelheden Benaming Snelheid m/s Windstil 0-0,2 Zwakke wind 0,3 3,3 Matige wind 3,4-7,9 Vrij krachtige wind 8,0 10,7 Krachtige wind 10,8 13,8 Harde wind 13,9 17,1 Stormachtig 17,2 20,7 De etmaalgemiddelde windsnelheden variëren gedurende het jaar flink en laten ook tussen de jaren een duidelijk verschil zien. In 2009 treedt er slechts enkele keren een etmaal gemiddelde windsnelheid hoger dan 10 m/s (krachtige wind) op. In 2010 gebeurt dit vaker. Echter buiten deze perioden van krachtige wind zijn de etmaalgemiddelde windsnelheden over het algemeen lager, zo rond de 4-5 m/s (matige wind). In 2011 zien we wederom meer momenten van krachtige wind gemiddelden over het etmaal. Opgemerkt wordt dat in dit jaar vlak voor de monsterperiode een etmaalgemiddelde windsnelheid van meer dan 12 m/s (krachtige wind) is opgetreden met hoogste uurgemiddelden rond de 17 m/s (stormachtig). Deze storm vindt plaats vlak na de zandsuppletie. In 2012 liggen de etmaalgemiddelde windsnelheden weer lager en komen de gemiddelde windsnelheden minder vaak boven de 10 m/s uit. Vlak voor de monsterperiode is zijn er verschillende stormachtige periodes. Echter etmaalgemiddeld is er sprake van een periode met gemiddeld matige wind. Net als het jaar daarna, in In 2013 zijn er echter meer periodes met krachtige wind en stormachtige wind per dag dan het jaar ervoor. Op 28 oktober en 5 december 2013 vinden er echte stormen plaats met hoogste uurgemiddelde windsnelheden van 20 en 18 m/s. Het laatste jaar, 2014 is een jaar van matige wind gemiddelde over het etmaal met een aantal dagen met periodes van krachtige wind. Opvalt dat in de periode voor de bemonstering een rustige periode is met zelfs geen dagen met krachtige wind. 3.2 Temperatuur In figuur 3.2 zijn de gemiddelde, minimale en maximale luchttemperatuur en de watertemperatuur weergeven voor de jaren 2009 tot en met
, Minimale luchttemperatuur (blauw) en maximale luchttemperatuur (rood) ter hoogte van Terschelling voor de jaren 2009 tot en met 2014 samen met de")
. In 2012 is deze periode korter dan in 2013, maar ligt de temperatuur wel lager.")
31 Figuur 3.2 Gemiddelde luchttemperatuur (zwart), Minimale luchttemperatuur (blauw) en maximale luchttemperatuur (rood) ter hoogte van Terschelling voor de jaren 2009 tot en met 2014 samen met de watertemperatuur ter hoogte van Schiermonnikoog. De blauwe lijn geeft de monsterperiode aan. De watertemperatuur bereikt in de zomer circa 19 graden in de bemonsterde jaren. In 2014 ligt dit zelfs een beetje hoger. De jaren 2012 en 2013 kennen beide een koude periode in het voorjaar (Figuur 3.2). In 2012 is deze periode korter dan in 2013, maar ligt de temperatuur wel lager. Wat betreft de bemonstering heeft deze niet altijd plaatsgevonden onder dezelfde omstandigheden qua temperatuur, wat mogelijk impact heeft om de bemonsterde macrobenthos. De belangrijkste verschillen tussen de jaren zijn weergegeven in tabel

32 Tabel 3.2 Overzicht van de maximale watertemperatuur, het moment van bemonstering en verdere opvallende afwijkingen in temperatuur, per jaar. Jaar Max watertemp Bemonstering C Maand C Maand Opvallend ,5 juli 19,5 juli Bemonstering in warmste periode juli 17 augustus Bemonstering tijdens afkoelen periode juli 17 augustus ,5 augustus 15 september Temperaturen tot -9 in februari. Bemonstering tijdens afkoelende periode juli 18 augustus Lange koude periode (tot -5) in het voorjaar (januari-april) juli 22 juli Bemonstering in warmste periode 3.3 Neerslag In figuur 3.3 zijn cumulatieve neerslag hoeveelheden per etmaal weergeven voor de jaren 2009 tot en met
33 Figuur 3.3 Neerslag per etmaal ter hoogte van Terschelling voor de jaren 2009 tot en met 2014.De blauwe lijn geeft de monsterperiode aan. De maximale neerslag per etmaal verschilt per jaar. In 2009 komt deze niet boven de 200 mm uit terwijl in de andere jaren maxima van mm gehaald worden. In de jaren 2010, 2011 en 2012 vindt er voorafgaand aan de monsterperiode neerslag van meer dan 240 mm per etmaal plaats. In 2013 en 2014 is dit niet het geval. In deze jaren is de neerslag over het gehele jaar ook lager. 26
. Het sediment wordt beschreven aan de hand van de gemiddelde korreldiameter (D50). Alle gebieden bestaan alle jaren gemiddeld uit fijn zand.")
34 4 Sedimentsamenstelling De mediane korrelgrootte hangt samen met de morfologie en het profiel van de vooroever, en wordt gezien als belangrijke abiotische parameter voor het functioneren van het ecosysteem (Speybroeck et al. 2006). Het sediment wordt beschreven aan de hand van de gemiddelde korreldiameter (D50). Alle gebieden bestaan alle jaren gemiddeld uit fijn zand. Binnen deze klasse treedt wel enige variatie op. De gemiddelde D50 per studiegebied varieert tussen de 158 en 208 μm met twee uitschieters in Ameland Uitstraling naar 121 μm in 2013 en 226 μm in De suppletie in Ameland Impact in 2011 lijkt weinig tot geen effect op de D50 te hebben (Figuur 4.1). Er is gesuppleerd met zand met een gemiddelde D50 van 153 μm (Arcadis 2009). Dit is dus iets fijner dan het oorspronkelijke materiaal. Dit komt vooral door een grotere fractie aan zeer fijn zand, gemiddeld 24%. Opgemerkt wordt dat de analyse van de monsters op de zandwinlocatie zijn uitgevoerd met behulp van zeven. De monsters op de suppletielocatie bij geanalyseerd met behulp van een Malvern. Hierdoor zijn deze gegevens niet een op een vergelijkbaar. En geven dus slechts een indicatie. Kijken we enkel naar de gegevens op de suppletielocatie (zie Tabel 4.1 waar een overzicht van de gemiddelde D50 per suppletiedieptezone, vooroever en zeewaarts van de suppletiedieptezone per studiegebied is weergegeven.) dan zien we dat de D50 van het sediment op de suppletielocatie varieert tussen de D50 tussen de 161 μm en de 181 μm. Figuur 4.1 Gemiddelde D50 per studiegebied per jaar. Het rode vak geeft het moment van suppleren in Ameland Impact weer. Bij een opname van het sediment vlak na aanleg van de suppletie in 2011 bleek de D50 juist iets grover, maar binnen de jaarlijkse variatie, te zijn dan in 2010 (van Egmond et al. 2010). Mogelijk dat tijdens de werkzaamheden de fijnere fractie reeds verdwenen was. Hier is verder geen onderzoek naar gedaan. 27
35 Vooroever Suppletie zone Zeewaarts suppletie zone Gebied Een effect van de suppletie is niet waarneembaar. Het verschil in D50 per jaar ten opzichte van het jaar 2010 is voor alle gebieden in de suppletiedieptezone en zeewaarts hiervan maximaal 15 μm. In de zone landwaarts van de suppletiedieptezone is dit verschil groter, vooral in Ameland Uitstraling (zie bijlage A voor de figuren). Tabel 4.1 Gemiddelde D50 per zone per studiegebied over de verschillende jaren. Het rode vak geeft het moment van suppleren op Ameland Impact weer. # 2009 (T0) 2010 (T0) 2011 (T1) 2012 (T2) 2013 (T3) 2014 (T4) Gem D50 Std # Gem D50 Std # Gem D50 Std # Gem D50 Std # Gem D50 Std # AI AM AU SR AI AM AU SR AI AM AU SR Ameland Schiermonnikoog Totaal Gem D50 Std 28
36 5 Resultaten macrobenthos 5.1 Soortenaccumulatiecurves De soortenaccumulatiecurve van alle gebieden (totaal) is weergegeven in figuur 5.1. De figuur laat zien dat er totaal over de jaren het hoogst aantal soorten is gevonden in Ameland Impact en het laagst aantal soorten op Schiermonnikoog. Doordat Ameland Midden minder bemonsterd is dan de overige gebieden, is het onduidelijk of het voldoende bemonsterd is en lijn asymptotisch wordt. Wel zijn er op Ameland Midden relatief veel soorten gevonden ten opzichte van het vaker bemonsterde Ameland Uitstraling en Schiermonnikoog met de relatief lage bemonstering die heeft plaatsgevonden. Figuur 5.1 Soortenaccumulatiecurve van alle monsters genomen over de jaren van alle gebieden (AI=Ameland Impact, AM=Ameland Midden, AU=Ameland Uitstraling, SR=Schiermonnikoog) Ameland Midden Figuur 5.2 laat de soortenaccumulatiecurve van Ameland Midden zien. Wanneer er meer wordt ingezoomd op het totaal van Ameland Midden, lijkt het totaal de asymptoot te benaderen. De grafiek laat zien dat in 2009 er waarschijnlijk te weinig monsters zijn genomen om de gemeenschap van Ameland Midden representatief weer te geven. In 2014 lijkt dit in mindere mate het geval te zijn. Figuur 5.2 Soortenaccumulatiecurve Ameland Midden. Het totaal aantal is gebaseerd op alle monsters, van alle jaren. Per jaar is het aantal gevonden soorten per monsteraantal weergegeven 29
37 5.1.2 Ameland Impact Op Ameland Impact was het hoogste aantal soorten gevonden over de jaren. Het gebied is dan ook het vaakst bemonsterd, en is gesuppleerd in Ook op Ameland Impact lijkt met name 2009 te weinig bemonsterd om de gemeenschap representatief weer te kunnen geven (Figuur 5.3), aangezien de lijn nog sterk stijgende is en er daardoor waarschijnlijk bij meer monsters nog meer soorten gevonden worden. Dit is ook het geval voor 2014, hoewel hier wel relatief veel soorten gevonden zijn. In 2010, 2012 en 2013 nadert de lijn een asymptoot, hierdoor lijkt het aantal genomen monsters aardig representatief voor de gemeenschap. In 2011 is de stijging van de lijn wel afgenomen, echter een asymptoot wordt nog niet genaderd, waardoor er waarschijnlijk te weinig is bemonsterd. Figuur 5.3 Soortenaccumulatiecurve Ameland Impact. Het totaal aantal is gebaseerd op alle monsters, van alle jaren. Per jaar is het aantal gevonden soorten per monsteraantal weergegeven Ameland Uitstraling Figuur 5.4 laat zien dat er vooral in 2013 waarschijnlijk voldoende monsters zijn genomen, aangezien er bijna geen nieuwe soorten gevonden worden bij het nemen van meer monsters. Dit is minder het geval voor 2010, 2011 en 2012, waarbij het aantal soorten dat gevonden wordt per monster nog steeds een stijging vertoond. Voor 2009 zijn er waarschijnlijk te weinig monsters genomen, aangezien de lijn nog sterk stijgt. De monsters van 2009 zijn dan ook niet representatief voor de gemeenschap. Figuur 5.4 Soortenaccumulatiecurve Ameland Uitstraling. Het totaal aantal is gebaseerd op alle monsters, van alle jaren. Per jaar is het aantal gevonden soorten per monsteraantal weergegeven 30
38 5.1.4 Schiermonnikoog De soortenaccumulatiecurve laat in 2012 een sterk stijgende lijn zien en het is daarom zeer onwaarschijnlijk dat de soorten van de genomen monsters representatief zijn voor de gehele gemeenschap in dit jaar. Ook in 2010 is de stijging in het aantal gevonden soorten per monster nog niet afgelopen, waardoor de monsters van dit jaar waarschijnlijk minder representatief zijn en 2012 laten een afname zien in het aantal nieuwe soorten dat gevonden wordt per extra monster, en genomen monsters zijn daardoor het meest representatief voor de gemeenschap in deze jaren ten opzichte van de overige jaren. In 2014 zijn relatief veel soorten gevonden voor het aantal monsters dat is genomen. Ook hier is de lijn nog stijgend en zijn er naar waarschijnlijkheid meer soorten in het gebied dan gevonden zijn in de monsters. Figuur 5.5 Soortenaccumulatiecurve Schiermonnikoog. Het totaal aantal is gebaseerd op alle monsters, van alle jaren. Per jaar is het aantal gevonden soorten per monsteraantal weergegeven 5.2 Vergelijking tussen studiegebieden Voor de vergelijking tussen de studiegebieden wordt er gekeken naar het gemiddelde per gebied voor biomassa, dichtheid en diversiteit. Dit is zowel gedaan voor de T0, waar de basisverschillen tussen de gebieden worden vergeleken, als voor alle bemonsterde jaren, waar op een grotere tijdschaal de gebieden worden vergeleken T0-vergelijking Biomassa Op moment T0-2010, voorafgaand aan de suppletie op Ameland Impact, laat de vergelijking van de soortensamenstelling op basis van biomassa zien dat vooral Ameland Impact en Ameland Uitstraling op elkaar lijken, verschillen zijn subtiel. Dit is niet verwonderlijk, gezien de gebieden vlak naast elkaar liggen. Schiermonnikoog wijkt significant in gemiddelde biomassa per monster (Tabel 5.1). Schiermonnikoog heeft een relatief hoge biomassa van de Amerikaanse zwaarschede (E. directus), en het ontbreekt aan de zeeklit (E. cordatum). Echter ook zonder deze soorten, wijkt Schiermonnikoog significant af in gemiddelde biomassa per monster. Dit wordt deels verklaard door de grote biomassa s borstelwormen (Nephtys) en van het zaagje (Donax vittatus) die op Ameland Impact en Ameland Uitstraling voorkomen ten opzichte van Schiermonnikoog. Doordat Ameland Impact en Ameland Uitstraling sterk op elkaar lijken tijdens de T0 2010, kan Ameland Impact na de suppletie goed vergeleken worden met Ameland Uitstraling. 31
 van de verschillende studiegebieden op T0 2010, excl E. directus en E. cordatum.")

39 Figuur 5.6 Gemiddelde biomassa per genomen monster (AI n=60; AU n=60, SR n=67) van de verschillende studiegebieden op T0 2010, incl E. directus en E. cordatum. Figuur 5.7 Gemiddelde biomassa per genomen monster (AI n=60; AU n=60, SR n=67) van de verschillende studiegebieden op T0 2010, excl E. directus en E. cordatum. Tabel 5.1 Uitkomsten ANOVA en Games-Howell post-hoc test voor biomassa. ANOVA + Games Howell post-hoc biomassa Biomassa incl E. directus + E. cordatum Biomassa excl E. directus + E. cordatum Paarsgewijze vergelijking p-waarde p-waarde AI AU 0,999 0,952 SR 0,000 0,000 AU AI 0,999 0,952 SR 0,000 0,000 SR AI 0,000 0,000 AU 0,000 0,000 32

40 Dichtheid De dichtheid op T laat zien dan Ameland Impact en Ameland Uitstraling veel overeenkomst vertonen. Schiermonnikoog laat meer afwijking zien met betrekking tot dichtheden van voorkomende soorten. Vooral de hoge dichtheden van borstelwormen zoals Magelona johnstoni op Ameland Impact en Ameland Uitstraling, zijn verklarend voor deze verschillen in gemiddelde dichtheid. Figuur 5.8 Gemiddelde dichtheid per genomen monster (AI n=60; AU n=60, SR n=67) van de verschillende studiegebieden op T Tabel 5.2 Uitkomsten paarsgewijze Games-Howell post-hoc test na ANOVA voor dichtheid. ANOVA + Games-Howell post-hoc dichtheid Paarsgewijze vergelijking p-waarde AI AU 0,998 SR 0,000 AU AI 0,998 SR 0,000 SR AI 0,000 AU 0,000 Diversiteit De Pielou s evenness van Schiermonnikoog ligt gemiddeld hoge dan voor Ameland. De diversiteit is gemiddeld het hoogst in Ameland Uitstraling, echter er zijn geen significante verschillen in diversiteit tussen de verschillende gebieden in T Tabel 5.3 Overzicht van diversiteitsparameters op van alle gebieden op moment T0 en de uitkomsten van de ANOVA en paarsgewijze Games-Howell post-hoc test. Gebied Monsters Richness Pielou's Shannon Paarsgewijze # soorten evenness Diversity (H') vergelijking AI AU 0,129 SR 0,999 AU AI 0,129 SR 0,096 SR AI 0,999 AU 0,096 33
 verantwoordelijk voor te zijn, doordat deze soort met een hoge gemiddelde biomassa aanwezig is in Ameland Midden en nagenoeg afwezig is op Schiermonnikoog.")
41 5.2.2 Gehele tijdsperiode Biomassa De gemiddelde biomassa van de studiegebieden ligt het hoogst in Ameland Midden en het laagst in Schiermonnikoog, dit verschil is significant (Figuur 5.9, Tabel 5.4). Hier lijkt vooral de zeeklit (E. cordatum) verantwoordelijk voor te zijn, doordat deze soort met een hoge gemiddelde biomassa aanwezig is in Ameland Midden en nagenoeg afwezig is op Schiermonnikoog. Tevens is de Amerikaanse zwaardschede (E. directus) een belangrijke soort voor de gemiddelde biomassa per monster, echter de biomassa van deze soort lijkt niet veel te verschillen tussen de gebieden. Figuur 5.9 Gemiddelde biomassa per gebied op basis van aantal monsters (AM n= 36; AI n= 285; AU n= 243, SR n= 172) inclusief E. directus en E. cordatum. Wanneer E. directus en E. cordatum buiten beschouwing worden gelaten (Figuur 5.10), zijn vooral het nonnetje (M. balthica) en de zandzager (Nephtys hombergii) belangrijke soorten voor de gemiddelde biomassa. Schiermonnikoog heeft ook een relatief grote bijdrage in biomassa van het gravend kniksprietkreeftje (Bathyporeia elegans) ten opzichte van de andere studiegebieden. Ameland impact lijkt op Ameland uitstraling wanneer E. directus en E. cordatum worden meegenomen. Zonder deze soorten komt Ameland Impact alleen overeen met Schiermonnikoog. Er zit relatief veel verschil tussen de gemiddelde biomassa tussen studiegebieden. Dit is niet vreemd, gezien het aantal monsters, het moment van monstername en de suppletiehistorie van de gebieden. 34

42 Figuur 5.10 Gemiddelde biomassa per gebied op basis van aantal monsters (AM n= 36; AI n= 285; AU n= 243, SR n= 172) exclusief E. directus en E. cordatum. Tabel 5.4 Games-Howell post-hoc biomassa Overzicht van de paarsgewijze vergelijkingen van gemiddelde totale biomassa per monster van de verschillende studiegebieden. Rode p-waarden geven significante verschillen aan, groene p- waarden geven niet significante verschillen aan. Biomassa incl E. directus + E. cordatum Biomassa excl E. directus + E. cordatum Paarsgewijze vergelijking p-waarde p-waarde AI AM AU SR AM 0,001 0,000 AU 0,469 0,026 SR 0,000 0,225 AI 0,001 0,000 AU 0,019 0,001 SR 0,000 0,000 AI 0,469 0,026 AM 0,019 0,001 SR 0,000 0,000 AI 0,000 0,225 AM 0,000 0,000 AU 0,000 0,000 Dichtheid Vooral de Amerikaanse zwaardschede (E. directus) en de borstelworm (Magelona johnstoni) zijn soorten die relatief veel bijdragen aan de totale dichtheid. Ameland Midden heeft een hogere gemiddelde dichtheid per monster dan de overige gebieden en heeft ten opzichte van de andere studiegebieden relatief hoge dichtheden witte dunschaal (Abra alba). Ameland Midden verschilt dan ook significant van de overige gebieden met betrekking tot de gemiddelde dichtheid (Tabel 5.5). Ameland Impact lijkt qua samenstelling en gemiddelde dichtheid sterk op Ameland Uitstraling, ondanks het feit dat Ameland Impact gesuppleerd is 35
 in relatief hoge dichtheden voor. Figuur 5.11 Gemiddelde dichtheid per genomen monster van de verschillende studiegebieden Tabel 5.")
43 en Ameland Uitstraling niet. Ameland Uitstraling en Schiermonnikoog verschillen onderling significant qua gemiddelde dichtheid. Wel komt in beide gebieden het vlokreeftje (Bathyporeia) in relatief hoge dichtheden voor. Figuur 5.11 Gemiddelde dichtheid per genomen monster van de verschillende studiegebieden Tabel 5.5 AI AM AU SR Uitkomsten ANOVA en Games-Howell post-hoc test op basis van gemiddelde dichtheid per monster per gebied ANOVA + Games-Howell post-hoc dichtheid Paarsgewijze vergelijking p-waarde AM 0,000 AU 0,259 SR 0,067 AI 0,000 AU 0,000 SR 0,000 AI 0,259 AM 0,000 SR 0,000 AI 0,067 AM 0,000 AU 0,000 36
44 Diversiteit Ameland Midden heeft een relatief hoge diversiteit ten opzichte van de andere studiegebieden (Tabel 5.6). De diversiteit van Ameland Impact, Ameland Uitstraling en Schiermonnikoog komen sterk overeen. Tabel 5.6 Gebied Overzicht van diversiteitsparameters en de uitkomsten van de ANOVA en paarsgewijze Games- Howell post-hoc test. Monsters Richness # soorten Pielou's evenness Shannon Diversity (H') AI AM AU SR Paarsgewijze vergelijking AM 0,011 AU 0,969 SR 0,996 AI 0,011 AU 0,005 SR 0,048 AI 0,969 AM 0,005 SR 0,935 AI 0,996 AM 0,048 AU 0,935 Uit de analyses voor biomassa, dichtheid en diversiteit blijkt dat vooral Ameland Impact, Ameland Uitstraling en Schiermonnikoog overeenkomsten vertonen. Ameland Midden wijkt meer af voor alle geanalyseerde parameters. Op Ameland Midden is slechts in 2 jaren bemonsterd. In de andere gebieden is minimaal 4 jaar bemonsterd. Dit kan het verschil tussen Ameland midden en de rest deels verklaren. Vooral in biomassa is er veel variatie, zowel inclusief als exclusief de relatief hoge biomassa soorten E. directus en E. cordatum. Vooral bij Ameland Uitstraling komt relatief veel M. balthica (nonnetje) voor. Hoewel Ameland Midden een relatief lage biomassa heeft en lage richness, kent het gebied een hogere diversiteit dan de overige gebieden. 5.3 Ontwikkeling per studiegebied Ameland Midden De ontwikkeling van Ameland midden over de jaren is geanalyseerd op basis van biomassa, dichtheid en diversiteit. Biomassa Voor Ameland Midden kan geen vergelijking tussen de jaren worden gemaakt in biomassa, doordat deze niet is bepaald in Figuur 5.12 toont daarom alleen de gemiddelde totale biomassa op Ameland Midden in
 inclusief E. directus en E. cordatum. B) exclusief E. directus en E. cordatum. Amerikaanse zwaardschede (E. directus) en de zeeklit (E.")
45 A) B) Figuur 5.12 Gemiddelde totale biomassa per monster per jaar (2014 n=36) op Ameland Midden weergegeven aan de hand van de 10 meest dominante soorten (en overige soorten gegroepeerd). A) inclusief E. directus en E. cordatum. B) exclusief E. directus en E. cordatum. Amerikaanse zwaardschede (E. directus) en de zeeklit (E. cordatum) maken een groot deel uit van de totale biomassa, doordat deze soorten relatief zwaar zijn per individu ten opzichte van andere soorten. Zonder deze soorten zijn vooral de zandzager (Nephtys hombergii) en het nonnetje (M. balthica) relatief hoog in biomassa. 38
. De jaren verschillen in gemiddelde totale dichtheid. Vooral de Amerikaanse zwaardschede (E.")
46 Dichtheid De gemiddelde dichtheid van het macrobenthos op de vooroever van Ameland Midden is in 2009 en 2014 bepaald (figuur 5.13). Suppletie Ameland Midden 2010 Figuur 5.13 Gemiddelde totale dichtheid per monster per jaar (2009 n=14, 2014 n=36). De jaren verschillen in gemiddelde totale dichtheid. Vooral de Amerikaanse zwaardschede (E. directus) is erg afgenomen tussen 2009 en Het verschil in gemiddelde dichtheid tussen de jaren is significant (p=0,001). De relatieve aanwezigheid van soorten lijkt meer gelijk verdeeld in 2014 dan in 2009, waar slechts 3 groepen dominant zijn. Diversiteit De diversiteit is significant toegenomen in 2014 ten opzichte van Dit bleek ook al uit figuur Ook de richness is toegenomen, wat mogelijk te wijten is aan het grotere aantal monsters dat genomen is in Tabel 5.7 Overzicht van diversiteitsparameters op Ameland Midden en de uitkomsten van de ANOVA en paarsgewijze Games-Howell post-hoc test. Jaar Monsters Richness Pielou's Shannon # soorten evenness Diversity (H') ,449 1, ,647 1,821 Paarsgewijze p-waarde diversity : 0, Ameland Impact De ontwikkeling van Ameland Impact over de jaren is geanalyseerd op basis van biomassa, dichtheid en diversiteit. Biomassa De gemiddelde totale biomassa per monster per jaar op Ameland Impact is weergegeven in figuur
 op Ameland Impact weergegeven aan de hand van de 10 meest dominante soorten (en overige")
 zijn dominante soorten voor de totale biomassa in alle jaren. Wanneer deze soorten buiten beschouwing worden gelaten zijn vooral het nonnetje (M.")
47 Suppletie 2011 A) Suppletie 2011 B) Figuur 5.14 Gemiddelde totale biomassa per monster per jaar (2010 n = 60, 2011 n=47, 2012 n=66, 2013 n=66, 2014 n=36) op Ameland Impact weergegeven aan de hand van de 10 meest dominante soorten (en overige soorten gegroepeerd). A) inclusief E. directus en E. cordatum. B) exclusief E. directus en E. cordatum. Vooral de Amerikaanse zwaardschede (E. directus) en de zeeklit (E. cordatum) zijn dominante soorten voor de totale biomassa in alle jaren. Wanneer deze soorten buiten beschouwing worden gelaten zijn vooral het nonnetje (M. balthica), de zandzager (Nephtys hombergii), de rechtgestreepte platschelp (Tellina fabula) en de borstelworm (Magelona johnstonii) dominant voor de totale gemiddelde biomassa. Opmerkelijk is de relatief hoge biomassa in 2011, een maand na suppleren van de vooroever op Ameland Impact. Er is in dit jaar vooral een hoge biomassa aan Amerikaanse zwaardschede (E. directus) aanwezig. Echter, ook de overige soorten verminderen nauwelijks in biomassa ten opzichte van In 40
48 2012 is er een significante afname in biomassa die grotendeels is toe te schrijven aan de afname in Amerikaanse zwaardschede (E. directus) dat jaar. Tevens vindt er in 2014 een significante toename plaats ten opzichte van de voorgaande jaren met uitzondering van 2010 in totale gemiddelde biomassa. Deze lijkt met name veroorzaakt te worden door een relatieve toename in de zandzager worm (Nephtys hombergii) en de het schelpdier witte dunschaal (Abra alba). Tabel 5.8 Games-Howell post-hoc biomassa Overzicht van de paarsgewijze uitkomsten van de Games-Howell post-hoc test op basis van de gemiddelde biomassa per monster per jaar. Biomassa incl E. directus + E. cordatum Biomassa excl E. directus + E. cordatum Paarsgewijze vergelijking p-waarde p-waarde ,702 0, ,123 0, ,106 0, ,009 0, ,702 0, ,012 0, ,978 1, ,497 0, ,123 0, ,012 0, ,000 0, ,000 0, ,106 0, ,978 1, ,000 0, ,590 0, ,009 0, ,497 0, ,000 0, ,590 0,001 41
.")
49 Dichtheid De gemiddelde totale dichtheid van macrobenthos op de vooroever van Ameland Impact is weergegeven in figuur Suppletie 2011 Figuur 5.15 Gemiddelde totale dichtheid van macrobenthos per jaar op de vooroever van Ameland Impact (2009 n= 17, 2010 n = 60, 2011 n=47, 2012 n=66, 2013 n=66, 2014 n=36). Over de jaren zijn vooral de Amerikaanse zwaardschede (E. directus) en de borstelworm (Magelona johnstoni) verantwoordelijk voor een grote bijdrage aan de totale dichtheid. Beide soorten verschillen sterk in dichtheid tussen de bemonsterde jaren. Opnieuw is de relatieve toename in dichtheid ten opzichte van 2010 opmerkelijk in 2011, een maand na suppleren. Statistisch verschilt de gemiddelde dichtheid op Ameland Impact na suppleren in 2011 niet van Ook is de zeer lage dichtheid in 2012 en de hoge dichtheid in 2009 opvallend. In 2014 lijkt er een grotere diversiteit aan soorten te zijn dan in eerdere jaren (Figuur 5.15). Dit zien we terug in de analyse van de diversiteit. Het bulldozerkreeftje (Urothoe poseidonis), de borstelworm (Spio martinensis) en het zeeklitschelpje (Tellimya ferruginosa) zijn bijvoorbeeld relatief toegenomen in dichtheid ten opzichte van eerdere jaren. 42
50 Tabel 5.9 Games-Howell post-hoc dichtheid Overzicht van de paarsgewijze uitkomsten van de Games-Howell post-hoc test op basis van gemiddelde dichtheid per monster per jaar. Paarsgewijze vergelijking Gemiddelde dichtheid per monster p-waarde , , , , , , , , , , , , , , , , , , , , , , , , , , , , , ,474 43
51 Diversiteit Uit de grafiek waarin de gemiddelde totale dichtheid en de dominante soorten van Ameland Impact door de jaren heen zijn weergegeven leek in 2014 de diversiteit te zijn toegenomen (Figuur 5.15). Dit wordt ook teruggevonden in de Shannon Diversity index die significant is toegenomen ten opzichte van eerdere jaren (Tabel 5.10). Ook de richness ligt hoger in 2014 ten opzichte van eerdere jaren, terwijl het aantal monsters lager ligt. Tabel 5.10 Jaar Overzicht van diversiteitsparameters en de uitkomsten van de ANOVA en paarsgewijze Games- Howell post-hoc test. Monsters Richness # soorten Pielou's evenness Shannon Diversity (H') paarsgewijze p-waarde diversity , , , , , , , , , , , , , , , , , , , , , , , , , , , , , ,000 44
 B) Figuur 5.")
. A) inclusief E. directus en E. cordatum. B) exclusief E. directus en E. cordatum. 45")
52 5.3.3 Ameland Uitstraling De ontwikkeling van Ameland Uitstraling over de jaren is geanalyseerd op basis van biomassa, dichtheid en diversiteit. Biomassa De gemiddelde totale biomassa van het macrobenthos van Ameland Uitstraling per jaar, is weergegeven in figuur 5.16-A en B. A) B) Figuur 5.16 Gemiddelde totale biomassa per monster per jaar (2010 n = 60, 2011 n=56, 2012 n=61, 2013 n=66) op Ameland Uitstraling weergegeven aan de hand van de 10 meest dominante soorten (en overige soorten gegroepeerd). A) inclusief E. directus en E. cordatum. B) exclusief E. directus en E. cordatum. 45
53 Figuur 5.16-A laat zien dat vooral de Amerikaanse zwaardschede (E. directus) en het nonnetje (M. balthica) een grote bijdrage leveren aan de gemiddelde biomassa. Met name in 2011 zijn er grote biomassa s Amerikaanse zwaardschede (E. directus) gevonden ten opzichte van de overige jaren. Voorafgaand aan de bemonstering van 2011 is er in Ameland Impact, liggende naast Ameland Uitstraling, gesuppleerd. Zonder de Amerikaanse zwaardschede (E. directus) en de zeeklit (E. cordatum) is het nonnetjes (M. balthica) de dominante soort voor biomassa, gevolgd door de zandzager (Nephtys hombergii) (Figuur 5.16-B). Er is geen significant verschil in biomassa tussen de jaren. Vooral in 2012 is de biomassa van het nonnetje (M. balthica) relatief hoog, zeker wanneer deze in de context van de relatief lage dichtheden in 2012 worden bekeken (Figuur 5.17). De bemonsterde individuen hadden dus een relatief hoge biomassa in Deze afwijking in 2012 zorgt echter niet voor een significant verschil tussen de jaren (Tabel 5.11). Tabel 5.11 Games-Howell post-hoc biomassa Overzicht van de paarsgewijze uitkomsten van de Games-Howell post-hoc test op basis van de gemiddelde biomassa per monster per jaar van Ameland Uitstraling. Biomassa incl E. directus + E. cordatum Biomassa excl E. directus + E. cordatum Paarsgewijze vergelijking p-waarde p-waarde ,009 0, ,756 0, ,000 0, ,009 0, ,004 0, ,841 0, ,756 0, ,004 0, ,000 1, ,000 0, ,841 0, ,000 1,000 46
.")
54 Dichtheid De gemiddelde dichtheid van het macrobenthos van AU is weergegeven in figuur Figuur 5.17 Gemiddelde totale dichtheid van macrobenthos per jaar op de vooroever van Ameland Uitstraling (2009 n= 6, 2010 n = 60, 2011 n=56, 2012 n=66, 2013 n=66). De jaarlijkse variatie in dichtheid van macrobenthos van de vooroever is groot op Ameland Uitstraling. De jaar tot jaar verschillen zijn significant op het jaar 2009 na. Vooral in 2012 ligt de gemiddelde dichtheid relatief laag. Dit werd ook gevonden voor Ameland Impact en Schiermonnikoog. De Amerikaanse zwaardschede (E. directus) en de borstelworm (Magelona johnstoni) zijn door de jaren heen de meest dominante soorten in dichtheid. De grote jaarlijkse variatie zorgt ervoor dat veel jaren significant van elkaar verschillen in gemiddelde dichtheid per monster (Tabel 5.12). 47
55 Tabel Games-Howell post-hoc biomassa Overzicht van de paarsgewijze uitkomsten van de Games-Howell post-hoc test op basis van gemiddelde dichtheid per monster per jaar van Ameland Uitstraling. Paarsgewijze vergelijking Gemiddelde dichtheid per monster p-waarde , , , , , , , , , , , , , , , , , , , ,000 48
56 Diversiteit De diversiteit en evenness zijn in 2013 significant afgenomen ten opzichte van eerdere jaren. Dit werd niet gevonden in de andere studiegebieden. Tabel 5.13 Jaar Overzicht van diversiteitsparameters en de uitkomsten van de ANOVA en paarsgewijze Games- Howell post-hoc test voor Ameland Uitstraling. Monsters Richness # soorten Pielou's evenness Shannon Diversity (H') paarsgewijze p-waarde diversity , , , , , , , , , , , , , , , , , , , ,001 49
 B) Figuur 5.")
. Het jaar 2012 bevat alleen diepgelegen monsters A) inclusief E. directus en E.")
57 5.3.4 Schiermonnikoog De ontwikkeling van Schiermonnikoog over de jaren is geanalyseerd op basis van biomassa, dichtheid en diversiteit. Biomassa De gemiddelde biomassa op Schiermonnikoog is weergegeven in figuur 5.18-A en B. A) B) Figuur 5.18 Gemiddelde totale biomassa per monster per jaar (2010 n = 60, 2011 n=60, 2012 n=16, 2013 n=36) op Schiermonnikoog weergegeven aan de hand van de 10 meest dominante soorten (en overige soorten gegroepeerd). Het jaar 2012 bevat alleen diepgelegen monsters A) inclusief E. directus en E. cordatum. B) exclusief E. directus en E. cordatum. 50
58 De Amerikaanse zwaardschede (E. directus) heeft ook op Schiermonnikoog een relatief grote bijdrage aan de totale gemiddelde biomassa. De zeeklit (E. cordatum) komt echter, contrasterend met de overige studiegebieden niet voor. Wel zijn het nonnetje (M. balthica) en de zandzager (Nephtys hombergii), zoals ook in overige gebieden dominante soorten voor de biomassa. Het nonnetje (M. balthica) neemt met een factor 4 toe in 2012 ten opzichte van Deze hoge biomassa van het nonnetje werd ook gevonden in Ameland Uitstraling in 2012 en in mindere mate ook in Ameland Impact. Deze hoge biomassa is opvallend, gezien de lage dichtheden die gevonden werden in In 2010 ligt de gemiddelde biomassa relatief laag, ook ten opzichte van de biomassa in Ameland Uitstraling en Ameland Impact in deze jaren. De hoge biomassa veroorzaakt door het nonnetje (M. balthica) in 2012 en lage biomassa in 2010, zorgt ervoor de gemiddelde biomassa van deze jaren significant afwijkt van de overige jaren (Tabel 5.14). Tabel 5.14 Overzicht van de paarsgewijze uitkomsten van de Games-Howell post-hoc test op basis van de gemiddelde biomassa per monster per jaar van Schiermonnikoog. Games-Howell post-hoc biomassa Biomassa incl E. directus + E. cordatum Biomassa excl E. directus + E. cordatum Paarsgewijze vergelijking p-waarde p-waarde ,000 0, ,000 0, ,000 0, ,000 0, ,000 0, ,001 0, ,970 0, ,000 0, ,000 0, ,001 0, ,002 0, ,001 0, ,000 0, ,970 0, ,002 0, ,983 0, ,000 0, ,000 0, ,001 0, ,983 0,998 Dichtheid De gemiddelde dichtheid van Schiermonnikoog varieert sterk tussen de jaren (Figuur 5.19). Ook de dominante soorten verschillen sterk in dichtheid en relatieve aanwezigheid tussen de jaren op Schiermonnikoog. Vooral in 2011 is de relatieve bijdrage van de Amerikaanse zwaardschede (E. directus) erg hoog, terwijl deze soort in overige jaren nauwelijks dominant aanwezig is voor dichtheid. Ook de borstelwormen Spio martinensis en Scololepis squamata en het gravend kniksprietkreeftje Bathyporeia elegans zijn soorten die in relatief hoge aantallen voorkomen op Schiermonnikoog ten opzichte van de studiegebieden van Ameland. De grote jaarlijkse variatie in dichtheid zorgt ervoor dat veel jaren significant van elkaar verschillen (Tabel 5.15). 51

59 Figuur 5.19 Gemiddelde totale dichtheid van macrobenthos per jaar op de vooroever van Schiermonnikoog (2010 n = 60, 2011 n=16, 2012 n=60, 2013 n=36). Tabel Games-Howell post-hoc biomassa Overzicht van de paarsgewijze uitkomsten van de Games-Howell post-hoc test op basis van gemiddelde dichtheid per monster per jaar van Schiermonnikoog. Paarsgewijze vergelijking Gemiddelde dichtheid per monster p-waarde , , , , , , , , , , , , , , , , , , , ,000 52
60 Diversiteit Zoals de overige parameters, verschil ook de diversiteit sterk tussen de jaren op Schiermonnikoog. Vooral in 2011 is de diversiteit en evenness erg laag (Tabel 5.16). In 2012 zijn er relatief weinig unieke soorten gevonden ten opzichte van de overige jaren, echter in dit jaar zijn er ook relatief weinig monsters genomen. Tabel 5.16 Jaar Overzicht van diversiteitsparameters en de uitkomsten van de ANOVA en paarsgewijze Games- Howell post-hoc test voor Schiermonnikoog. Monsters Richness # soorten Pielou's evenness Shannon Diversity (H') paarsgewijze p-waarde diversity , , , , , , , , , , , , , , , , , , , , Suppletiedieptezone De suppletie wordt enkel in een deel van het studiegebied aangelegd. In deze paragraaf wordt de ontwikkeling per studiegebied in enkel de suppletiedieptezone weergegeven voor biomassa, dichtheid en diversiteit. Daarnaast wordt de verspreiding van biomassa en dichtheid voor Ameland en Schiermonnikoog (bijlage C.1 biomassa en C.2 dichtheid) weergegeven. Biomassa Gelijk aan de analyses voor het gehele bied zijn in de suppletie zone dezelfde soorten grotendeels dominant over de jaren in de drie verschillende studiegebieden. Vooral Ameland Uitstraling en Ameland Impact komen overeen met betrekking tot biomassa en soortensamenstelling, binnen dezelfde jaren, met uitzondering van 2012 waarin op Ameland Uitstraling een erg hoge biomassa van het nonnetje (M. balthica) bemonsterd is. 53
61 Schiermonnikoog kent een andere jaar tot jaar ontwikkeling in biomassa dan de studiegebieden van Ameland. Tussen 2010 en 2011 nam de biomassa (inclusief E. directus en E. cordatum, figuur 5.20-A, C en E) toe in het suppletiegebied van het wel-gesuppleerde Ameland Impact en het nietgesuppleerde Ameland uitstraling. Zonder deze dominante soorten voor de biomassa is er op Ameland Impact echter een kleine afname in biomassa tussen de periode voor de suppletie en 1 maand na de suppletie, ten opzichte van de stabiele biomassa in Ameland Uitstraling (Figuur 5.20-D en E). De ontwikkeling van de samenstelling van soorten door de jaren heen het zaagje (Donax vittatus) en de witte dunschaal (Abra alba) sterk afnemen in biomassa zowel in de suppletiedieptezone van Ameland Uitstraling als van Ameland Impact. In alle studiegebieden komt de borstelworm (Nepthys cirrosa) in 2013 (25 maanden sinds suppleren) in relatief hoge biomassa s voor. Ameland Uitstraling heeft in 2011 en 2012 een relatief hoge biomassa de tere platschelp (Tellina tenuis) ten opzichte van Ameland Impact en Schiermonnikoog, waar deze soort niet dominant voorkomt. Suppletie 2011 Suppletie 2011 A) B) C) D) 54
, Ameland Uitstraling (B en E) en Schiermonnikoog (C en F), weergegeven aan de hand van het aantal maanden sinds")

62 E) F) Figuur 5.20 Gemiddelde biomassa van de suppletiedieptezone van Ameland Impact (A en D), Ameland Uitstraling (B en E) en Schiermonnikoog (C en F), weergegeven aan de hand van het aantal maanden sinds de suppletie op Ameland Impact in Inclusief (A, B en C) en exclusief (D, E en F) E. directus en E. cordatum. De totale gemiddelde biomassa in de suppletiedieptezone is dus niet afgenomen als gevolg van de suppletie een maand na de suppletie. Echter, verspreidingsfiguren van de biomassa in het gesuppleerde Ameland Impact en het uitstralingsgebied Ameland Uitstraling, laten wel een lokale afname van biomassa zien aan de zeewaartse zijde van de suppletie. De jaren na 2011 keren de biomassa s op deze monsterlocaties langzaam terug Figuur
63 A) B) 56
64 C) D) 57
65 E) Figuur 5.21 Verspreiding van de gemiddelde biomassa per monsterpunt, gekoppeld aan de coördinaten van het monster. De grootte van het monsterpunt geeft de gemiddelde biomassa van het monsterpunt weer. De rode lijn geeft de suppletiezone weer in Ameland Impact. Rechts van het gesuppleerde gebied ligt Ameland Uitstraling. A) 2010, B) 2011, C) 2012, D) Het oranje vak in B (2011) geeft de monsters met een gemiddelde biomassa <0,01 weer. De verspreidingsfiguur laat zien dat er in 2011 na de suppletie op Ameland Impact en een stuk van Ameland Uitstraling een aantal monsterpunten in de suppletiezone is waar de biomassa nagenoeg 0 is. Er zijn in 2011 ook een aantal monsterpunten in de suppletiedieptezone van Ameland impact waar de biomassa relatief hoog is. Dit kan verklaren waarom de afname in grafiek met de gemiddelde biomassa op de suppletiedieptezone niet evident is (Figuur 5.20). De verspreidingsfiguur laat daarnaast zien dat hoewel 2012 over het algemeen een jaar was met relatief lage biomassa s, dit niet uitgesproken lager was in de suppletiedieptezone van Ameland Impact tov Ameland Uitstraling. In 2013 lijkt de biomassa van alle monsterpunten in de suppletiedieptezone van Ameland Impact en Ameland Uitstraling op elkaar en zijn er geen opvallend lage biomassa monsterpunten meer aanwezig. Dichtheid Ook op basis van dichtheid is er meer gelijkenis tussen de suppletiedieptezone van Ameland Uitstraling en Ameland Impact met betrekking tot de jaarlijkse variatie en soortensamenstelling. Dit blijkt ook uit een ANOSIM (Tabel 5.17) waaruit blijkt dat hoewel alle gebieden significant verschillen, de r-waarde voor Ameland Impact en Uitstraling erg laag is en deze gebieden meer overeenkomen met elkaar dan met Schiermonnikoog. Ten opzichte van Ameland Uitstraling heeft Ameland Impact meer van de borstelworm Spio martinensis. Schiermonnikoog wijkt meer af van de suppletiedieptezones van Ameland, zowel 58
 is opvallend in 2011 (0 maanden sinds de suppletie op Ameland Impact).")
66 in ontwikkeling over de jaren als in soortensamenstelling. Vooral de piek in de Amerikaanse zwaardschede (E. directus) is opvallend in 2011 (0 maanden sinds de suppletie op Ameland Impact). 25 en 36 maanden na de suppletie lijkt ook de diversiteit van soorten hoger te liggen op Schiermonnikoog. Suppletie 2011 Suppletie 2011 A) D) B) E) C) F) 59
, Ameland Uitstraling (B) en Schiermonnikoog (C), weergegeven aan de hand van het aantal maanden sinds de suppletie op Ameland")

67 Figuur 5.22 Gemiddelde dichtheid van de suppletiedieptezone van Ameland Impact (A), Ameland Uitstraling (B) en Schiermonnikoog (C), weergegeven aan de hand van het aantal maanden sinds de suppletie op Ameland Impact in Inclusief E. directus en E. cordatum. Wanneer er naar de korte-termijn impact van de suppletie gekeken wordt op Ameland Impact, vergeleken met Ameland Uitstraling, is er een maand na de suppletie zelfs een kleine toename in dichtheid. De borstelworm Magelona mirabilis neemt af direct na de suppletie, echter dit is ook het geval voor Ameland Uitstraling waar niet gesuppleerd is. In 2012 (14 maanden sinds de suppletie) is een jaar met een lage dichtheid. Dit werd voor de meeste soorten (excl. M. balthica) ook gevonden voor de biomassa. De opportunistische borstelworm Magelona johnstoni komt direct na de suppletie wel in lagere dichtheden voor. In 2014 lijkt de diversiteit voor verschillende soorten hoger te liggen doordat meer soorten in gelijke aantallen voorkomen en er niet 1 of 2 dominant aanwezig zijn. Dit is ook het beeld in Schiermonnikoog. Ameland Uitstraling is niet bemonsterd in Aan de hand van de dichtheid is er geen duidelijk effect van de suppletie, zowel op relatief korte als op wat langere termijn te aanschouwen op de gemiddelde dichtheid per monster, doordat dichtheden niet afnemen in het suppletiegebied van Ameland Impact. Ook veranderingen in de soortensamenstelling komen overeen met het niet gesuppleerde Ameland Uitstraling. Verspreidingsfiguren van de gemiddelde dichtheid per monsterpunt, gekoppeld aan de coördinaten van het monsterpunt, laten een afname in dichtheid zien specifiek in de suppletiedieptezone van Ameland Impact en een deel van Ameland Uitstraling in 2011 na aanleg van de suppletie (Figuur 5.23). In 2012 zijn over het gehele studiegebied van Ameland (Ameland Impact en Ameland Uitstraling) maar ook in Schiermonnikoog (niet weergegeven) de dichtheden laag, onafhankelijk van de suppletiedieptezone. In 2013 liggen de dichtheden in de suppletiedieptezone van Ameland Impact ongeveer in dezelfde range van de dichtheden van omliggende monsterpunten op dezelfde diepte. 60
68 A) B) 61
69 C) D) 62
70 E) Figuur 5.23 Verspreiding van de gemiddelde dichtheid per monsterpunt, gekoppeld aan de coördinaten van het monster. De grootte van het monsterpunt geeft de gemiddelde dichtheid van het monsterpunt weer. De rode lijn geeft de suppletiezone weer in Ameland Impact. Rechts van het gesuppleerde gebied ligt Ameland Uitstraling. A) 2010, B) 2011, C) 2012, D) Het oranje vak in B (2011) geeft de monsters met een gemiddelde dichtheid beneden 10 weer. Een MDS plot bevestigt het beeld uit de verspreidingsfiguren van de gemiddelde dichtheid per monsterpunt. De MDS plot van de gemiddelde dichtheid van de monsters laat zien dat er na de suppletie in 2011, 5 monsters (omcirkeld in figuur 5.24) waren die afweken van de overige monsters uit de suppletiedieptezone van 2011 (alle gebieden, donkerblauw). De afwijkende monsters liggen allen aan het meest zeewaarts gelegen gedeelte van de suppletie, dus op het deel waar de golfwerking het sterkst is na de suppletie. Het jaar erop zijn er nog steeds een tweetal monsterpunten van AI die afwijken ten opzichte van de monsters van overige gebieden in deze zone. Twee jaar na de suppletie is dit voor slechts 1 monster nog het geval. Deze MDS plot laat zien dat enkele monsters in de suppletiedieptezone sterk anders zijn ten opzichte van de overige monsters in de suppletiedieptezone dat jaar. Er is dus wel een impact van de suppletie zichtbaar in Ameland Impact op de gemiddelde dichtheid per monster, echter deze is slechts zichtbaar in een aantal monsterpunten. Jaar tot jaar variatie in alle gebieden is groot wat ook blijkt uit een MDS plot op basis van dichtheid (Figuur 5.25). 63
71 Figuur 5.24 MDS plot op basis van dichtheid van de suppletiedieptezone verschillende maanden na het suppleren (kleuren) en van verschillende studiegebieden (AI= Ameland Impact, AU= Ameland uitstraling, SR= Schiermonnikoog). Monsters van AI die afwijken van de overige monsters zijn omcirkeld, deze omcircelde monsters liggen aan de meest zeewaarts gelegen rand van de suppletiezone. De kleuren geven het aantal maanden na de suppletie op Ameland Impact weer, de afkortingen voor gebieden zijn naast de monsterpunten weergegeven. Figuur 5.25 MDS plot op basis van dichtheid van de suppletiedieptezone van verschillende studiegebieden (kleuren) en aantal maanden na suppleren op Ameland Impact in 2011 (genummerd: -10, 1, 14, 25, 36). Tabel 5.17 Uitkomsten Two-way crossed ANOSIM (factors gebied en jaar). Een hoge R-waarde (dichtbij 1) betekent dat er een grote scheiding is tussen de factoren (gebied), een lage R-waarde (dichtbij 0) betekent dat er weinig verschil is tussen de factoren. Gebied R-waarde P-waarde (Two-way crossed ANOSIM) Ameland Impact Ameland Uitstraling 0,066 0,001 Ameland Impact Schiermonnikoog 0,54 0,001 Ameland Uitstraling - Schiermonnikoog 0,58 0,001 64
72 Diversiteit De Shannon-Wiener index voor diversiteit en Pielou s evenness index zijn weergegeven in de tijd, zodat de ontwikkeling van verschillende gebieden in de suppletiedieptezone meer inzichtelijk is. Zowel Shannon-Wieners diversity index als Pielou s evenness index laten hetzelfde patroon zien per studiegebied, wat niet vreemd is omdat evenness ook wordt meegenomen in de Shannon-Wieners diversity index. De diversiteit en gemiddelde verdeling van soorten lag redelijk hoog in In 2011 namen beide parameters iets af in alle gebieden. Op Ameland Impact nam de index iets meer af dan op Ameland Uitstraling en iets minder dan op Schiermonnikoog na de suppletie die plaatsvond 1 maand voor het monitoren. Echter in 2012 nam de diversiteit en evenness wel meer toe dan op Ameland Uitstraling. Over de tijd zit er veel variatie in de diversiteit en evenness en een duidelijk effect van de suppletie is niet zichtbaar. Suppletie 2011 Suppletie 2011 A) Figuur 5.26 B) A) Shannon-Wiener diversiteitsindex van de studiegebieden Ameland Impact (AI), Ameland Uitstraling (AU) en Schiermonnikoog (SR) over de tijd weergegeven. B) Pielou s evenness index van de studiegebieden Ameland Impact (AI), Ameland Uitstraling (AU) en Schiermonnikoog (SR) over de tijd weergegeven. 5.5 Losse soorten uitgelicht Doordat er voor bepaalde soorten met name een effect wordt verwacht van de suppletie, zijn er een aantal soorten uitgelicht waar nader op wordt ingezoomd. Deze soorten zijn: Macoma balthica (nonnetje): belangrijke vertegenwoordiger van de langlevende soorten Nephtys hombergii (zandzager): opportunistische aas-etende soort die mogelijk op korte termijn een voordeelpositie kan genieten. Echinocardium cordatum (zeeklit): verplaatst zich makkelijk en heeft een grote invloed op de totale biomassa. Magelona johnstoni (borstelworm): opportunistische soort die mogelijk op korte termijn een voordeelpositie kan genieten vanwege de snelle rekrutering. Van een deze individuele soorten is zowel de dichtheid, biomassa als de biomassa/dichtheid (een maat voor de biomassa per individu) weergegeven. Hierbij is een onderscheid gemaakt tussen voorkomen in de suppletiedieptezone, de ondiepe zone en de zeewaartse zone. 65

73 Verder is de verspreiding in dichtheid per soort in Ameland en Schiermonnikoog (in bijlage C) weergegeven Macoma balthica De gemiddelde biomassa is nagenoeg gelijk gebleven voor Ameland Impact kort na de suppletie, terwijl er op Ameland Uitstraling en Schiermonnikoog een kleine toename zichtbaar is. De biomassa/dichtheid laat zien dat zowel in 2010 als in 2011 de aanwezige individuen relatief groot zijn ten opzichte van de andere studiegebieden, maar dat er in 2011 wel kleinere individuen aanwezig zijn dan in Kleine individuen kunnen zich beter verplaatsen dan grote individuen van het nonnetje (M. balthica) en dit kan verklaren dat er zo kort na de suppletie weer redelijk grote aantallen aanwezig zijn met een relatief lage biomassa. In 2012 zijn de aanwezige individuen gemiddeld erg groot, vooral op Ameland Uitstraling. Dit is opmerkelijk omdat er in dit jaar lage dichtheden en biomassa s van soorten gevonden is, met uitzondering van het nonnetje (M. balthica). Suppletie 2011 AI Suppletie 2011 AI A) B) Suppletie 2011 AI C) Figuur 5.27 Gemiddelde dichtheid (A), biomassa (B) en dichtheid per individu (C) van M. balthica. Het nonnetje (M. balthica) is op Ameland Impact na de suppletie van 2011 gemiddeld toegenomen in dichtheid in het gehele gebied, maar ook in de suppletiedieptezone (Figuur 5.27). Deze toename in dichtheid werd ook gevonden op Schiermonnikoog en Ameland Uitstraling. Op Ameland Uitstraling ligt de dichtheid van het nonnetje (M. balthica) gemiddeld 66
74 hoger in alle jaren, terwijl Schiermonnikoog en Ameland Impact meer op elkaar lijken. Het gesuppleerde Ameland Impact laat geen duidelijke afwijking zien met betrekking tot ontwikkeling van de dichtheid van het nonnetje (M. balthica). Op Ameland Uitstraling is deze toename nog sterker dan in de overige gebieden. Wanneer gemiddelde dichtheden van monsterpunten in verspreidingsfiguren wordt weergegeven, wordt het effect van de suppletie duidelijker (Figuur 5.28). A) B) 67
75 C) D) 68
76 Figuur 5.28 Verspreiding van de gemiddelde dichtheid per monsterpunt van M. balthica, gekoppeld aan de coördinaten van het monster. De grootte van het monsterpunt geeft de gemiddelde dichtheid van deze soort op het monsterpunt weer. De rode lijn geeft de suppletiezone weer in Ameland Impact. Rechts van het gesuppleerde gebied ligt Ameland Uitstraling. A) 2010, B) 2011, C) 2012, D) De oranje box in C (2011) geeft opvallende punten weer in de suppletiedieptezone van Ameland Impact. De paarse box dient als vergelijking met de oranje box in 2011 voor de dichtheden van M. balthica in het naastgelegen Ameland Uitstraling op de suppletiedieptezone, waar niet gesuppleerd is in E) De verspreidingsfiguren van de gemiddelde dichtheid per monsterpunt laten zien dat 2010 een jaar was met lage gemiddelde dichtheden op de vooroever van heel Ameland. Opvallend is dat in het meest oostelijke deel er een licht hogere dichtheid gevonden is. Dit blijft zichtbaar in de jaren hierna. In 2011 lijken dichtheden gemiddeld gezien te zijn toegenomen, met uitzondering van de individuen op de suppletiedieptezone van Ameland Impact en een deel van Ameland Uitstraling. Enkele relatief grote dichtheden op de rand van de suppletiedieptezone van Ameland Impact zouden kunnen verklaren waarom in de grafiek van de gemiddelde dichtheid in de suppletiedieptezone wel toeneemt (Figuur 5.27). In 2013 komen de dichtheden van de gehele suppletiedieptezone van Ameland weer meer met elkaar overeen. Over het algemeen laten de verschillende parameters zien dat M. balthica na de suppletie in nagenoeg gelijke gemiddelde dichtheden en biomassa s voorkomt, zowel in de suppletiedieptezone als daarbuiten. Er zijn echter wel lokaal effecten te zien in de dichtheid in de suppletiedieptezone van Ameland Impact en een deel van Ameland Uitstraling. Ten opzichte van het overige deel van Ameland Uitstraling blijft Ameland Impact in de suppletiedieptezone achter in de toename in dichtheid. Na ongeveer 2 jaar komen de dichtheden van de gehele suppletiedieptezone van Ameland weer meer met elkaar overeen. 69

 op Ameland Impact en Ameland Uitstraling laat direct na de suppletie een afname zien, die meer uitgesproken is voor Ameland Impact met name in")
77 5.5.2 Magelona johnstoni De opportunistische borstelworm Magelona johnstoni is direct na de suppletie afgenomen in gemiddelde dichtheid per monster (Figuur 5.29). Deze afname is echter zowel zichtbaar in Ameland Impact en Ameland Uitstraling. Op Ameland Impact is de afname meer uitgesproken dan voor Ameland Uitstraling, vooral in de suppletiedieptezone. Schiermonnikoog laat een toename zien in dichtheid. Ook de biomassa van de borstelworm (Magelona johnstoni) op Ameland Impact en Ameland Uitstraling laat direct na de suppletie een afname zien, die meer uitgesproken is voor Ameland Impact met name in de ondiepe zone. De grafiek die de biomassa/dichtheid toont (Figuur 5.29-C), laat zien dat de aanwezige individuen direct na de suppletie een stuk groter zijn op Ameland Impact, vooral in de ondiepe zone. Grote soorten komen vooral in de ondiepe zone voor in alle gebieden over alle jaren. Dit is vooral uitgesproken in 2011, in de overige jaren ligt de gemiddelde biomassa/dichtheid meer gelijk. Op termijn verschilt de dichtheid, biomassa en biomassa/dichtheid niet veel tussen Ameland Impact en Ameland Uitstraling. Op Schiermonnikoog liggen de waarden gemiddeld lager. Suppletie 2011 AI A) Suppletie 2011 B) AI Suppletie 2011 AI C) Figuur 5.29 Gemiddelde dichtheid (A), biomassa (B) en dichtheid per individu (C) van Magelona johnstoni. Doordat er op Ameland Impact direct na de suppletie een afname in dichtheid is ten opzichte van de overige gebieden, zou dit mogelijk door de suppletie veroorzaakt kunnen zijn. In de verspreidingsfiguren van Figuur 5.30 wordt dit beeld bevestigd. In 2011 is er een relatieve 70
 op Ameland toegenomen tov 2012.")
78 afname van Magelona johnstoni dichtheden op de suppletiedieptezone van Ameland Impact en enigszins op dezelfde diepte in het naastgelegen uitstralingsgebied. In 2012 zijn alle dichtheden van de worm op Ameland vergelijkbaar laag. In 2013 is de borstelworm (Magelona johnstoni) op Ameland toegenomen tov A) B) 71
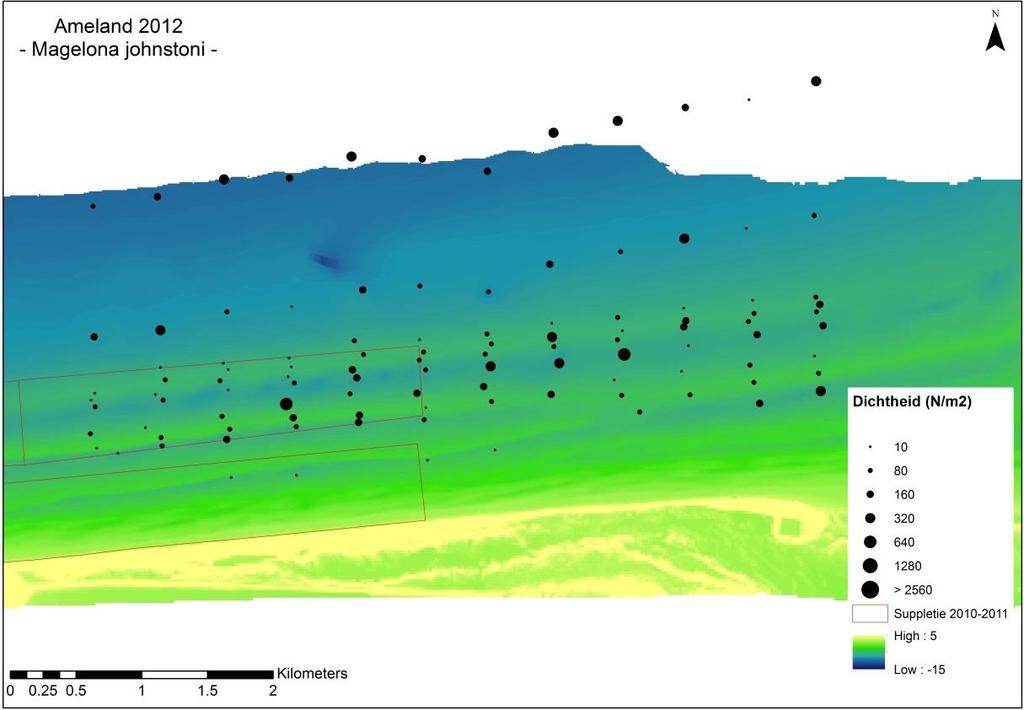
79 C) D) 72

80 Figuur 5.30 Verspreiding van de gemiddelde dichtheid per monsterpunt van Magelona johnstoni, gekoppeld aan de coördinaten van het monster. De grootte van het monsterpunt geeft de gemiddelde dichtheid van deze soort op het monsterpunt weer. De rode lijn geeft de suppletiezone weer in Ameland Impact. Rechts van het gesuppleerde gebied ligt Ameland Uitstraling. A) 2010, B) 2011, C) 2012, D) Oranje pijlen in 2011 (C) geven opvallende dichtheden aan. E) Nepthys hombergii De zandzager Nepthys hombergii is een maand na de suppletie op Ameland Impact (2011) afgenomen op Ameland Impact. Echter deze afname is ook zichtbaar op Ameland Uitstraling. Schiermonnikoog ondergaat een toename in de zandzager (Nepthys hombergii). De afname op Ameland Impact is meer uitgesproken dan op Ameland Uitstraling, echter dit verschil is beperkt. Het is daardoor moeilijk om de afname in Ameland Impact te koppelen aan de suppletie. De biomassa volgt hetzelfde patroon tussen 2010 en 2011 als de dichtheid, alleen is het verschil in afname tussen Ameland Impact en Ameland Uitstraling kleiner. Over het algemeen zijn er geen duidelijke afwijkingen van de zandzager (Nephtys hombergii) op Ameland Impact ten opzichte van Ameland Uitstraling en Schiermonnikoog over de jaren, die te koppelen zijn aan de suppletie. Deze bevindingen worden bevestigd door de verspreidingsfiguren van de dichtheid van Nephtys hombergii (Figuur 5.32). 73
")


81 A) B) Suppletie 2011 AI Suppletie 2011 AI C) Figuur 5.31 Gemiddelde dichtheid (A), biomassa (B) en dichtheid per individu (C) van Nephtys hombergii. 74
82 A) B) 75
83 C) D) 76
84 Figuur 5.32 Verspreiding van de gemiddelde dichtheid per monsterpunt van Nephtys hombergii, gekoppeld aan de coördinaten van het monster. De grootte van het monsterpunt geeft de gemiddelde dichtheid van deze soort op het monsterpunt weer. De rode lijn geeft de suppletiezone weer in Ameland Impact. Rechts van het gesuppleerde gebied ligt Ameland Uitstraling. A) 2010, B) 2011, C) 2012, D) Oranje box in 2011 (C) geeft de overeenkomst in dichtheden op de suppletiedieptezone weer van Nephtys hombergii in zowel Ameland Impact als Ameland Uitstraling. E) 77


85 5.5.4 Echinocardium cordatum De zeeklit (E. cordatum) had een relatief hoge dichtheid en biomassa in Ameland Impact in Beide parameters zijn afgenomen voor de soort in Deze afname is minder uitgesproken voor Ameland Uitstraling voor dichtheid en is afwezig voor biomassa. In de jaren later na de suppletie ( ) laat het gesuppleerde Ameland Impact geen verdere afwijkingen zien. Het is onduidelijk of de suppletie een effect heeft op de soort vanwege de afname die ook gevonden werd in op Ameland Uitstraling. De afname in de suppletiezone is echter wel meer uitgesproken dan in de diepere zone op Ameland Impact. De verspreidingsfiguren bevestigen dit beeld. Ameland Uitstraling laat een sterkere afname zien van E. cordatum dichtheden (Figuur 5.34). Het is moeilijk om conclusies te trekken op basis van de suppletie, doordat deze soort over het algemeen meer patchy voorkomt en vaak in de meer diepe gebieden. Suppletie 2011 AI A) B) Suppletie 2011 AI Suppletie 2011 AI C) Figuur 5.33 Gemiddelde dichtheid (A), biomassa (B) en dichtheid per individu (C) van E. cordatum. 78
86 A) B) 79
87 C) D) 80
88 Figuur 5.34 Verspreiding van de gemiddelde dichtheid per monsterpunt van E. cordatum, gekoppeld aan de coördinaten van het monster. De grootte van het monsterpunt geeft de gemiddelde dichtheid van deze soort op het monsterpunt weer. De rode lijn geeft de suppletiezone weer in Ameland Impact. Rechts van het gesuppleerde gebied ligt Ameland Uitstraling. A) 2010, B) 2011, C) 2012, D) Oranje pijlen in 2011 (C) geven opvallende dichtheden aan. E) 81
89 82
90 6 Conclusies 6.1 Gebiedsvergelijkingen H0-1: Alle studiegebieden zijn gelijk in dichtheid, biomassa en diversiteit onderling over de hele periode In de T0-situatie in 2010 komen Ameland Impact en Ameland Uitstraling duidelijk overeen voor dichtheid en biomassa. Schiermonnikoog wijkt meer af. Qua diversiteit en richness zijn alle gebieden vergelijkbaar in de T0-situatie. Wanneer alle gebieden vergeleken zouden worden op grote tijdschaal, kan verwacht worden dat de gebieden zullen verschillen vanwege hun verschillende historie en verloop binnen de periode van vier jaar monsteren (gesuppleerd/niet-gesuppleerd). Dit wordt ook gevonden in de analyses. Ameland Impact lijkt vooral op Ameland Uitstraling en in mindere mate op Schiermonnikoog. Ameland Midden wijkt meer af van de andere studiegebieden doordat het een gemiddeld hogere biomassa en dichtheid heeft. Opgemerkt wordt dat dit gebied minder vaak bemonsterd is dan de overige studiegebieden. Over de jaren is de jaarvariatie in ieder studiegebied zeer duidelijk aanwezig. De ontwikkeling van de dichtheid is enigszins vergelijkbaar tussen Ameland Uitstraling en Ameland Impact. Voor de gemiddelde biomassa wordt dit niet waargenomen. Deze verschillen worden vaak veroorzaakt door verschillen in specifieke soorten zoals het nonnetje in Schiermonnikoog wijkt over het algemeen meer af van de studiegebieden van Ameland. Dominante soorten voor dichtheid verschillen tussen Ameland en Schiermonnikoog. Zo komen het gravend kniksprietkreeftje (Bathyporeia elegans) en de borstelwormen Spio martinensis en Scololepis squamata relatief veel voor op Schiermonnikoog ten opzichte van Ameland, waar juist E. cordatum relatief veel voorkomt. Gezien de afwijkende geschiedenis (nooit gesuppleerd) en tevens de afwijkende gradiënt van de vooroever (minder steil) is het goed verklaarbaar dat de vooroever van Schiermonnikoog zich anders gedraagt. Aan de hand van deze bevindingen kan de H0-1 verworpen worden. 6.2 Ontwikkeling per studiegebied H0-2: De afzonderlijke studiegebieden komen overeen tussen de jaren in dichtheid, biomassa en diversiteit. Ameland Midden is slechts 2 jaar bemonsterd (2009; 2014) en in 2009 enkel voor dichtheid. De dichtheid en diversiteit verschillen significant tussen deze jaren. Ameland Impact is in alle jaren bemonsterd voor biomassa en dichtheid. Voor biomassa laat een vergelijking tussen de jaren zien dat er in 2014 een significante toename heeft plaatsgevonden ten opzichte van de voorgaande jaren met uitzondering van 2010, het jaar voorafgaand aan de suppletie. De hogere biomassa in 2014 zit vooral in een relatieve toename in de zandzager worm (Nephtys hombergii) en de het schelpdier witte dunschaal 83
91 (Abra alba). Met betrekking tot de dichtheid is het opmerkelijk dat de totale gemiddelde dichtheid in 2011 na suppleren niet significant is afgenomen ten opzichte van Het jaar 2012 is een jaar waarin de dichtheid significant lager is ten opzichte van alle andere jaren. Ondanks dat de dichtheid in 2014 niet significant verschillend is ten opzichte van de andere jaren is de diversiteit in 2014 wel significant hoger. Dit wordt veroorzaakt doordat de verdeling van de gemiddelde dichtheid per soort gelijkmatiger is. Daarnaast zijn er in 2014 ook meer soorten gevonden. In Ameland Uitstraling is geen significant verschil gevonden in biomassa tussen de jaren. Hoewel er een relatief hoge biomassa van het Nonnetje is waargenomen in 2012 zorgt dit niet voor een significant verschil in de gemiddelde totale biomassa tussen de jaren. Met betrekking tot de dichtheid is de jaarlijkse variatie in het macrobenthos van de vooroever groot. De jaar tot jaar verschillen zijn significant op het jaar 2009 na. Vooral in 2012 ligt de gemiddelde dichtheid relatief laag. De Amerikaanse zwaardschede (E. directus) en de borstelworm (Magelona johnstoni) zijn door de jaren heen de meest dominante soorten in dichtheid. De diversiteit en evenness zijn in 2013 significant afgenomen ten opzichte van eerdere jaren. In Schiermonnikoog is in 2010 de gemiddelde biomassa significant lager dan in de overige jaren. In 2012 is de biomassa juist significant hoger. Dit wordt veroorzaakt door het nonnetje (M. balthica) welke in hoge biomassa s aanwezig is. De gemiddelde dichtheid en de dominante soorten varieren sterk tussen de jaren. De grote jaarlijkse variatie in dichtheid zorgt ervoor dat veel jaren significant van elkaar verschillen. Vooral in 2011 is de relatieve bijdrage van de Amerikaanse zwaardschede (E. directus) erg hoog, terwijl deze soort in overige jaren nauwelijks dominant aanwezig is voor dichtheid. Ook de diversiteit verschilt sterk tussen de jaren op Schiermonnikoog. Vooral 2011 is significant lager dan de andere jaren. Op basis van deze bevindingen kan geconcludeerd worden dat de H0-2 verworpen kan worden en dat binnen de studiegebieden er variatie is tussen de jaren. 6.3 Suppletiedieptezone H0-3: Er is geen verschil in de ontwikkeling van het macrobenthos in de suppletiedieptezone in het gesuppleerde Ameland Impact, ten opzichte van het niet-gesuppleerde Ameland Uitstraling en Schiermonnikoog. Wanneer er wordt ingezoomd op de suppletiedieptezone van de verschillende studiegebieden (met uitzondering van Ameland Midden, omdat deze op een ander moment gesuppleerd is) lijken de gebiedsontwikkelingen van Ameland Impact en Ameland Uitstraling wat betreft gemiddelde totale biomassa en dichtheid per monster op elkaar. De totale gemiddelde biomassa en dichtheid in de suppletiedieptezone is nauwelijks afgenomen als gevolg van de suppletie een maand na de suppletie. De verspreidingsfiguren (biomassa en dichtheid) laten echter zien dat er zeer lokaal wel veranderingen optreden in de meest zeewaarts gelegen zone in de suppletiedieptezone van Ameland Impact. Ter hoogte van de verandering in de suppletiediepte zone van Ameland Impact wordt aangrenzend in Ameland Uitstraling een vergelijkbaar effect waargenomen. Dit effect is zichtbaar direct na de suppletie. Dit wordt bevestigd door de MDS plot op basis van de gemiddelde dichtheid van monsterpunten in de suppletiedieptezone, welke een afwijking 84
92 laat zien in de soortensamenstelling van de buitenste monsterpunten in de suppletiedieptezone van Ameland Impact, direct na de suppletie. De jaren hierna, beginnen deze monsterpunten weer steeds meer op de overige monsterpunten in de suppletiedieptezone te lijken. Deze resultaten duiden erop dat er in ieder geval zeer lokaal een effect is van de suppletie in het suppletiegebied direct na suppleren. Dit effect lijkt gemiddeld gesproken na 2 jaar niet meer te onderscheiden. Aan de hand van deze bevindingen kan de H0-3 verworpen worden, omdat er wel een verschil is in de ontwikkeling van het macrobenthos op de suppletiedieptezone van Ameland Impact en deels in het niet-gesuppleerde Ameland Uitstraling. 6.4 Losse soorten uitgelicht Doordat er op losse soorten mogelijk een impact verwacht wordt van de suppletie, die tekenend kan zijn voor een soortgroep, zijn er een aantal losse soorten geanalyseerd over de jaren in verschillende zones van de vooroever. De gemiddelde dichtheid en biomassa van het nonnetje (M. balthica) laten een maand na de suppletie op Ameland geen verschil zien, zowel in de suppletiedieptezone als daarbuiten. Echter de verspreidingskaarten laten zien dat het nonnetje sterk afneemt in dichtheid direct na de suppletie op enkele monsterlocaties in de suppletiedieptezone in Ameland Impact en deels in Ameland Uitstraling. Ongeveer twee jaar na de suppletie lijken de dichtheden in de suppletiedieptezone van Ameland Impact weer op de omliggende monsterpunten op dezelfde diepte. Deze bevindingen komen overeen met de tijdreeksanalyse op gegevens van aangespoelde bodemdieren langs de Nederlandse kust (Gmelig Meyling et al. 2009). Hier werd een hersteltijd van 1 tot 3 jaar voor tweekleppigen afgeleid. De gemiddelde dichtheid en biomassa van opportunistische borstelworm Magelona johnstoni is direct na de suppletie afgenomen. Deze afname is zowel zichtbaar in Ameland Impact als Ameland Uitstraling. Op Ameland Impact is de afname meer uitgesproken dan voor Ameland Uitstraling, vooral in de suppletiedieptezone. Schiermonnikoog laat juist een toename zien in dichtheid, hoewel de gemiddelde waarden lager liggen dan op Ameland. Op basis van de verspreidingskaarten laat de borstelworm Magelona johnstoni laat een afname in dichtheid op de suppletiedieptezone zien in de zeewaarts gelegen zone van Ameland Impact en deels van Ameland Uitstraling. De afname betreft vooral kleinere individuen. In 2012 zijn alle dichtheden van de worm op Ameland vergelijkbaar laag. In 2013 is de borstelworm (Magelona johnstoni) op Ameland toegenomen ten opzichte van De zandzager (Nephtys hombergii) en de zeeklit (E. cordatum) ondergaan geen herkenbare effecten op de korte of lange termijn als gevolg van de suppletie. Door de losse soorten uit te lichten, kan er geconcludeerd worden dat geanalyseerde soorten verschillend reageren op de vooroeversuppletie. Sommige soorten ervaren meer impact dan anderen. Over het algemeen vindt er rekolonisatie plaats van de soorten die een impact ervaren, in een periode van ongeveer 2-3 jaar. 85
93 86
94 7 Doorvertaling naar de suppletiepraktijk De natuurlijke zandige kust van Nederland wordt met behulp van zandsuppleties veilig gehouden en kan ruimte bieden aan diverse gebruiksfuncties. Hoewel zandsuppleties worden gezien als noodzakelijk voor het handhaven van de functies van de kust bestaat er de zorg dat de huidige suppleties van invloed zijn op het functioneren van het ecosysteem van de ondiepe kustzone. Deze zorg stond aan de basis van het convenant dat Rijkswaterstaat met de vier natuurbeschermingsorganisaties (Stichting de Noordzee, de Waddenvereniging, Stichting Duinbehoud en de Vogelbescherming) heeft opgesteld. De doelstelling vanuit het convenant luidt: Het verkrijgen van meer inzicht ten aanzien van de invloed van de uitvoering van zandsuppleties op de natuurwaarden en mogelijkheden van het optimaliseren van het suppletieprogramma en de uitvoering daarvan met het behoud en de ontwikkeling van de natuurwaarden. Jaarlijks wordt er langs de hele Nederlandse kust ongeveer 12 Mm 3 zand aangebracht. Het benodigde zand wordt gewonnen buiten de doorgetrokken -20 meter diepte lijn en langs de kust op de bodem aangebracht tussen de -5 en -8 meter diepte. De hoeveelheid die op een locatie wordt aangebracht wordt bepaald door middel van een jaarlijkse toetsing van de basiskustlijn. Bij het ontwerpen van een suppletie worden grofweg de volgende ontwerprichtlijnen gehanteerd (van der Spek et al. 2007): Volume: Frequentie: Kustdwarse positie: Afhankelijk van de jaarlijkse toetsing van de BKL. Varieert van elke 2 tot 4 jaar afhankelijk van de locatie. Bij voorkeur onderwater in de actieve zone (vanaf -8 meter en ondieper) van het kustsysteem. Eventueel langs een geulwand of buiten de buitenste brekerbank op circa -5 tot -8 meter diepte. Anders op het strand. Sedimentsamenstelling: Zoveel mogelijk gelijk aan het oorspronkelijke sediment op de locatie. Moment van suppleren: Buiten het broedseizoen. Op basis van de casestudie op Ameland en Schiermonnikoog naar de effecten van de suppletie op Ameland midden in is de volgende kennis opgedaan ten behoeve van deze ontwerprichtlijnen. Met moet er rekening mee houden dat het hier een studie van een enkele suppletie in het Waddengebied betreft. 7.1 Suppletievolume De suppletie bij Ameland-midden was een grote suppletie ten opzichte van de andere suppleties uitgevoerd in het suppletieprogramma (zie ook Figuur 1.2). De suppletie is mede hierdoor zeer gefaseerd aangelegd waardoor de impact van dit volume verspreid is in ruimte en tijd. De eventuele positieve en/of negatieve uitwerking van het suppletievolume of de gefaseerde uitvoering hiervan op het bodemleven is in deze studie niet nader onderzocht. er is geen nieuwe kennis die aanleiding geeft om de huidige richtlijn voor het suppletievolume te wijzigen. 87
95 7.2 Suppletiefrequentie In het studiegebied Ameland-Impact en het naastgelegen Ameland Uitstraling is ter hoogte van de suppletielocatie zeer lokaal een afname waargenomen in de gemiddelde biomassa en dichtheid per monsterpunt welke lijkt te worden veroorzaakt door de suppletie. Dit effect wordt na twee circa jaar niet meer duidelijk waargenomen. Kijken we naar specifieke soorten verschilt deze periode. Zo laat het nonnetje pas na twee jaar gemiddelde dichtheden zien die vergelijkbaar zijn aan dichtheden voorafgaand aan de suppletie. Terwijl dit bij de borstel worm Magelona johnstoni al na een jaar is en de zandzager Nepthys hombergii laat helemaal geen effect van de suppletie zien. deze kennis geeft geen aanleiding om de huidige suppletiefrequentie te wijzigen. 7.3 Sedimentsamenstelling Er is gesuppleerd met zand met een gemiddelde D50 welke binnen de marge van de D50 van het oorspronkelijk aanwezige zand lag. Op basis van bodemmonsters in het suppletiegebied wordt er over de jaren een verschil in gemiddelde D50 waargenomen van 15 μm. Deze variatie heeft geen effect op de aanwezige bodemfauna. deze kennis geeft geen aanleiding om de huidige richtlijn voor de sedimentsamenstelling te wijzigen. 7.4 Kustdwarse positie De vooroever van Ameland kent een dubbel banken patroon. Dit in tegenstelling tot Schiermonnikoog. De vooroever is permanent in beweging en laat jaarlijkse netto hoogte verschillen zien van meer dan 2 meter. Dit geeft aan dat het hier een dynamische zone van de kust betreft (Holzhauer et al. 2014). De suppletie in Ameland-Impact is over de meest zeewaarts gelegen bank aangelegd. Na aanleg is op de suppletielocatie direct een nieuwe bank 3 de ontstaan die de eerdere banken naar de kust drukt (Vermaas et al. 2013). Het effect van de positie van de suppletie op het macrobenthos van de vooroever van Ameland is in deze studie niet nader onderzocht. Wel is gebleken dat op de positie van deze specifieke suppletie, vanaf 5 meter en dieper over de meest zeewaarts gelegen bank, geen onderscheidende bodemdiergemeenschappen zijn waargenomen op de toppen, hellingen en troggen van de banken. Ondanks dat er o.a. volgens Janssen et al. (2008) aanwijzingen zijn dat er een verhoogde biodiversiteit in troggen zou moeten zijn, verklaard door lagere stroomsnelheden, hoger slibgehalte, minder dynamiek in de troggen. Deze bevinding is tijdens dit onderzoek niet vastgesteld. Dit wil echter niet direct zeggen dat er zeer lokaal geen verhoogde diversiteit in de troggen aanwezig kan zijn. Het bemonsteren van lokale clusters met hogere diversiteit aan bodemdieren vraagt gebiedsdekkende monitortechnieken. Het bemonsteren via bodemhappen is hiervoor onvoldoende geschikt. Tijdens de bemonsteringen bij Ameland is geconstateerd dat over het algemeen het vooral de afstand tot de kust en de diepte zijn, die de faunasamenstelling bepalen waarbij het niet uit maakt of dat de monsters op de top van een bank zijn genomen of in een trog (Holzhauer et al. 2014). Kijken we naar de volledige gradiënt van laagwater tot buitenste brekerbank, dan is er sprake van een grove driedeling. Rond laagwater lijkt de bodemdiersamenstelling en het vóórkomen van bodemdieren erg op de bodemdiergemeenschappen van het lage strand en vormen dus een duidelijke overgang tussen het strand en de vooroever. Vervolgens vormt de bankenzone een geheel wat betreft 88
96 bodemdiersamenstelling. De derde zone wordt gevormd door de bodemdiermonsters net buiten de bankenzone tot circa 12 meter diepte. Een meer detail opdeling in morfologische eenheden lijkt dus niet nodig. Voor het suppletieonderzoek betekent dit dat het volledige suppletiediepte zone samen geanalyseerd kan worden, hetgeen de power van de resultaten verhoogt. deze kennis geeft geen aanleiding om de huidige richtlijn voor de kustdwarse positie te wijzigen. 7.5 Moment van suppleren De suppletie Ameland-midden is zeer gefaseerd uitgevoerd en heeft bijna een heel jaar in beslag genomen. Hierdoor was het niet mogelijk om een effect van het moment van suppleren waar te nemen. Wel is voorafgaand aan de suppletie een bemonstering uitgevoerd naar de aanwezige schelpdieren. De resultaten hiervan gaven geen aanleiding tot het niet uitvoeren van de suppletie (Goudswaard et al. 2009). Er is geen nieuwe kennis die aanleiding geeft om de huidige richtlijn voor het moment van suppleren te wijzigen. 89
97 90


98 8 Discussie De rol van jaarlijkse variatie De resultaten van de analyses laten zien dat er een impact van de suppletie is in de suppletiedieptezone voor zowel gemiddelde totale biomassa als dichtheid. Deze impact neemt af over een periode van ongeveer 2 jaar, wanneer de bodemdiergemeenschap weer sterk lijkt op die van Ameland Uitstraling. De jaar tot jaar variatie in de samenstelling, dichtheid en biomassa van het macrobenthos is zeer groot, groter dan het effect van de suppletie. Dit bemoeilijkt het direct herkennen van de effecten van een suppletie en zorgt ervoor dat in sommige grafieken in deze rapportage de effecten van de suppletie niet zichtbaar zijn. Echter wanneer er voor de jaarlijkse variatie wordt gecorrigeerd is er wel een effect te herkennen op de locatie van de daadwerkelijke suppletie. De verspreidingsfiguren (Figuur 5.21 en Figuur 5.23) en de MDS plot (Figuur 5.24) gaven reeds dit inzicht. Naderhand is dit nogmaals getest met behulp van een PCA analyse waarbij zoveel mogelijk gecorrigeerd is voor de jaarvariatie (Figuur 8.1). Figuur 8.1 Effecten van de suppletie bij Ameland Impact (2011). In deze analyse is de structuur van de gemeenschap geanalyseerd met een PCA (principal component analysis) op basis van dichtheid van de soorten met een frequentie > 5%. De PCA toont de dominante assen van variatie in de fauna. De diagrammen rechts geven de resultaten van deze analyse, apart per jaar voor de grafische duidelijkheid (maar de analyse is voor alle jaren samen gebeurd). De dominante gradiënt correleert met de diepte, en beide assen zijn min of meer kwadratisch gecorreleerd. In 2011 vallen een aantal stations duidelijk buiten de verwachte range. Dat blijkt ook uit de kaartjes, waar de waarde van de stations op de eerste PCA-as zijn geïnterpoleerd en als kaart gepresenteerd. Rond (en vooral zeewaarts) van de suppletie valt in 2011 een vlek op van een fauna die meer verwant is aan de fauna van het strand, dan aan de fauna die normaal op deze diepte wordt gevonden. 91
99 Een ruimtelijke weergave van de PCA analyse (links in figuur 8.1) laat de monsterpunten van Ameland Impact zien voor de verschillende jaren. De kleuren geven de ligging van het monsterpunt op de eerste PCA-as weer. Het ruimtelijke beeld laat zien dat de monsterpunten sterk correleren met diepte (verloop van de kleuren). Opgemerkt wordt dat diepte waarschijnlijk geen causale factor is maar eerder een proxy voor de onderliggende fysische omstandigheden die verklarend zijn voor de bodemdiersamenstelling. Tevens is het suppletie-effect in 2011 duidelijk zichtbaar, zeer lokaal in de buitenste monsters van de suppletiedieptezone. De PCA toont hiermee aan dat er een effect van de suppletie aanwezig is die onafhankelijk is van de jaarvariatie. Verklaring lokaal effect Opvallend is dat er enkel een zeer lokaal effect lijkt op te treden in Ameland Impact maar ook in een deel van het naastgelegen Ameland Uitstraling. Hiervoor kunnen verschillende verklaringen gevonden worden. De eerste verklaring heeft mogelijk te maken met de uitvoeringspraktijk. Aangezien deze suppletie zeer gefaseerd is uitgevoerd is het mogelijk dat dit deel van Ameland Impact als allerlaatste is gesuppleerd. Terwijl het meer westelijk gelegen deel eerder in de tijd is gesuppleerd. Een tweede verklaring heeft mogelijk te maken met de fysische omstandigheden aan de meest zeewaarts gelegen zijde van de suppletie. Doordat op de meest zeewaarts gelegen zijde van de suppletie de golfwerking waarschijnlijk versterkt is, kan dit verklaren waarom er juist op de buitenste monsterpunten van de suppletie een grotere impact zichtbaar is. De omstandigheden zijn hier waarschijnlijk dusdanig bar geworden, dat de bodemdiersamenstelling direct na de suppletie van 2011 meer lijkt op die van meer ondiepe gebieden, die over het algemeen minder rijk zijn (kleurverloop 2011, Figuur 8.1). Dit effect is zeer sterk verminderd na een jaar en helemaal weg na 2 jaar. Fysische en meteorologische impact De grote natuurlijke dynamiek in een gebied zoals de vooroever van Ameland kan het snelle herstel van de bodemdiergemeenschap verklaren. De vooroever is gedurende het jaar sterk in beweging (Vermaas et al. 2013). Daarnaast blijkt uit de meteorologische gegevens dat er veel verschillen tussen de jaren zijn in temperatuur, regenval en stormen. Zowel gemiddelden als extreme situaties kunnen het macrobenthos beïnvloeden, bijvoorbeeld door relatief veel input van zoetwater wat zoutstress zou kunnen veroorzaken bij macrobenthische soorten (Telesh et al. 2013), stormen die de verdeling van het sediment en turbiditeit beïnvloeden (Birchenough et al. 2011), maar ook extreme temperaturen die de levenscyclus van soorten kan beïnvloeden (Hiscock et al. 2004). Zo heeft het vlak voor de bemonstering in 2012 gestormd. Het kan goed zijn dat dit van invloed is geweest op de resultaten. Echter geeft de vergelijking met Schiermonnikoog hier helaas geen uitsluitsel omdat Schiermonnikoog in te veel aspecten afwijkt van Ameland. Aangepast aan de dynamische omgeving Doordat de bodemdiergemeenschap in de natuurlijke situatie veel dynamiek kent, is het waarschijnlijk dat de samenstelling van de soorten hierop is afgestemd en is aangepast. Echter, de effecten van de suppletie, met name in de buitenste rand van de suppletie zijn klaarblijkelijk van dusdanige omvang ten opzichte van de natuurlijke dynamiek, dat ze op korte termijn voor een andere soortensamenstelling zorgen meer lijkend op de soortensamenstelling van de ondiepe strata waar minder soorten voorkomen. 92
100 Is Schiermonnikoog wel geschikt als referentiegebied? Uit de analyses bleek dat het referentiegebied Schiermonnikoog zich anders ontwikkelde dan Ameland Impact of Ameland Uitstraling. Hoewel Schiermonnikoog eenzelfde bankenpatroon kent als Ameland, is de vooroever flauwer en is het nooit gesuppleerd. De omstandigheden van dit gebied verschillen daardoor van die van de vooroever van Ameland welke vanwege de voortdurende erosie wel regelmatig gesuppleerd moet worden. Vanwege de grote jaarlijkse variatie en de lokale verschillen is het van belang zijn om zowel meerdere referentiegebieden te volgen als de ontwikkeling in het impact gebied. Ontwikkeling binnen een jaar In deze studie is enkel gekeken naar de netto jaarlijkse ontwikkeling. Wat er gedurende het jaar gebeurd is niet apart bekeken. Het kan nuttig zijn om de ontwikkeling binnen een jaar te bekijken om zo de waargenomen jaar op jaar verschillen correct te kunnen interpreteren. De variabiliteit in de omgevingsfactoren die de bodemgemeenschap tevens beïnvloeden en daarmee het beeld over de effecten van suppleties kunnen vertroebelen, kunnen hiermee tevens meer inzichtelijk gemaakt worden. Opschalen naar een landelijk beeld Deze studie betreft een enkele suppletie in het Waddengebied. Om te toetsen dezelfde uitkomsten gelden voor de gehele Nederlandse kust, zal ook hier studie naar gedaan moeten worden. Dit kan op verschillende manieren, 1) in de vorm van het opvolgen van een suppletie op een nieuwe locatie, 2) door enkele vaste raaien langs de kust te kiezen waarlangs de bodemdierontwikkeling wordt opgevolgd ongeacht of er een suppletie wordt uitgevoerd of niet, of 3) door elke suppletie langs de nederlandse kust kort op te volgen met enkele raaien aan monsters. Sediment is belangrijk De korrelgrootteverdeling is één van de belangrijkste omgevingsvariabelen voor de samenstelling van de bodemdierengemeenschap die zowel kustdwars als kustlangs varieert langs de Nederlandse kust. De hydrodynamiek leidt tot sortering van het sediment. Over het algemeen zien we langs de Nederlandse kust een kustdwars profiel waarin de mediane korrelgrootte het hoogst is in de brandingszone en afneemt richting de diepere vooroever. Korrelgrootte is bepalend voor de stabiliteit van gangen in het sediment, de doorgraafbaarheid van het sediment, en de permeabiliteit. Deze laatste is mogelijk de belangrijkste parameter. Permeabiliteit bepaalt hoe sterk het interstitiële water meestroomt met het bovenstaande water. In sterk permeabele sedimenten is deze doorstroming groot, waardoor deze sediment maar weinig fijn materiaal kunnen vasthouden. Zij zijn zeer arm aan koolstof, die echter van hoge kwaliteit is. Wanneer de doorstroming beperkter is, hoopt meer organisch detritus (van lagere kwaliteit) in het sediment op. Dat is gunstig voor deposit feeders die leven op deze voedselbron. Over het algemeen treedt er een verarming (in biomassa, dichtheid, aantal soorten) op naarmate het sediment grover wordt. Bij de suppletie Ameland-midden is er met sediment gesuppleerd overeenkomstig met het oorspronkelijke sediment wat een effect bepaling bemoeilijkt. Daarnaast is deze suppletie op Ameland-midden zeer gefaseerd uitgevoerd wat mogelijk het sorteringsproces heeft geholpen. Om te toetsen in hoeverre een suppletie de bodemsamenstelling 93
101 daadwerkelijk veranderd is studie nodig waarbij de ontwikkeling van het sediment in beeld wordt gebracht en er gewerkt wordt met verschillende sedimentsamenstellingen. Het ecosysteem verandert kustdwars Elke zone van de kust is anders. In het geval van de suppletie op Ameland-midden is de suppletie in een zeer actieve zone van de kust aangelegd. Wanneer een suppletie in een rustigere zone wordt aangelegd, bijvoorbeeld op dieper water, zal wederom naar de effecten op het bodemleven gekeken moeten worden. Op basis van deze enkele case is het nog niet mogelijk om een relatie tussen de dynamiek van de kust en de status van het daar aanwezig bodemleven in te schatten. Hiervoor zal de dynamiek en het bodemleven op meerdere plekken kustlangs en kustdwars onderzocht moeten worden. De rol van een case-studie voor begrip van de Nederlandse kust De studie bij Ameland heeft ons veel geleerd over het herstel van het bodemleven in de bankenzone na een relatief grote vooroeversuppletie. Tevens heeft het ons inzicht gegeven in de aanwezige bodemdieren en zonering over het kustprofiel. Om te toetsen of dezelfde uitkomsten gelden voor de gehele Nederlandse kust, zal ook hier studie naar gedaan moeten worden. De vergelijking met het toch zeer nabij gelegen Schiermonnikoog onderschrijft dit. Studie op de schaal van de gehele Nederlandse kust kan op verschillende manieren worden aangepakt, 1) in de vorm van het opvolgen van een suppletie op een nieuwe locatie, 2) door enkele vaste raaien langs de kust te kiezen waarlangs de bodemdierontwikkeling wordt opgevolgd ongeacht of er een suppletie wordt uitgevoerd of niet, of 3) door elke suppletie langs de Nederlandse kust kort op te volgen met enkele raaien aan monsters. Naast de opschaling naar de gehele Nederlandse kust zullen ook de cumulatieve effecten van jarenlang suppleren langs de kust op de samenstelling van de bodemdieren in ogenschouw genomen moeten worden. Lange termijn kustdekkend onderzoek zou hier meer inzicht in kunnen bieden. 94
102 9 Literatuur Arcadis (2009). Grondonderzoek zandwingebieden, Arcadis. Birchenough, S. N. R., S. Degrear, H. Reiss, A. Borja, U. Braeckman, J. Craeymeersch, I. De Mesel, F. Kerckhof, I. Kröncke, N. Mieszkowska, S. Parra, M. Rabaut, A. Schröder, C. Van Colen, G. Van Hoey, M. Vincx en K. Wätjen (2011). Responses of marine benthos to climate change ICES Cooperative Research Report No. 310 ICES. Faasse, M., K. Lock en T. Vanagt (2013). Labrapport benthos Ameland. Najaar Oostende, ecoast. Faasse, M. en T. Vanagt (2014). Labrapport Strand- en vooroeverbenthos Ameland. Najaar Oostend, ecoast. Gmelig Meyling, A. W. en R. H. de Bruyne (2009). Onder het zand beland. Effecten van strand- en onderwatersuppleties op het macro- en epibenthos van de nabije kustzone onderzocht met behulp van Systematisch Strandonderzoek (pilotstudie) Periode Bennebroek, Stichting anemoon/ Metridium. Goudswaard, K., J. Kesteloo, C. van Zweeden en T. Vanagt (2009). Kwantitatieve bemonstering in het zandsuppletiegebied Ameland in 2009 op de aanwezigheid van schelpdierbanken. Yerseke, IMARES & Grontmij. Heusinkveld, J. (2013). Monsterverslag Ameland subtidaal September Monstername macrobenthos tbv onderzoek naar effecten van kustsuppleties. Groningen. Heusinkveld, J. (2014). Monsterverslag Ameland subtidaal September Monstername macrobenthos tbv onderzoek naar effecten van kustsuppleties. Groningen. Heusinkveld, J. en T. Vanagt (2011). Monsterverslag veldcampagne ecologie Ameland Belgie, Oostende, The Fieldwork Company, ecoast. Hiscock, K., A. Southward, I. Tittley en S. Hawkinds (2004). "Effect of changing temperature on benthic marine life in Britain and Ireland " Aquatic Conservation: Marine and Freshwater Ecosystems 14(4): Holzhauer, H., J. Reinders, L. M. Vonhögen, A. De Backer en J. Heusinkveld (2012). Meetplan veldcampagne Ameland 2012, Deltares. Holzhauer, H., B. van der Valk, J. van Dalfsen, M. Baptist en G. Janssen (2009). Ecologisch gericht suppleren, nu en in de toekomst. Het ontwerp meerjarenplan voor monitoring en (toepassingsgericht) onderzoek. Delft, Deltares, IMARES & VU Amsterdam. Holzhauer, H., T. Vanagt, K. Lock, M. C. van Oeveren, A. De Backer, K. Hostens, J. van Dalfsen en J. Reinders (2014). Ecologische effecten suppletie Ameland Interim rapportage ihkv KPP B&O Kust Ecologie. Delft, Deltares. Ibanez, M. E. (2014). Sediment analysis Ameland Delft, Deltares. Ibanez, M. E. en M. A. de Lucas Pardo (2014). Sedimentanalyse Ameland studie. Delft, Deltares. 95
103 Janssen, G. M., H. Kleef, S. Mulder en P. Tydeman (2008). "Pilot assessment of depth related distribution of macrofauna in surf zone along Dutch coast and its implications for coastal management." Marine Ecology 29(1): Leewis, L. (2011). Afweging aanpak sedimentanalyses 2011 t.b.v. de monitoring suppletie Ameland 2010/2011. Amsterdam, Grontmij. Leewis, L., J. Heusinkveld en E. C. Verduin (2012). Veldcampagne Ameland 2011 Veldverslag. Amsterdam, Grontmij Nederland B.V. Ministerie van Infrastructuur en Milieu (2015). Kustlijnkaarten Lelystad, Rijkswaterstaat. Speybroeck, J., D. Bonte, W. Courtens, T. Gheskiere, P. Grootaert, J. P. Maelfait, M. Mathys, S. Provoost, K. Sabbe, E. W. M. Stienen, V. Van Lancker, M. Vincx en S. Degraer (2006). "Beach nourishment: An ecologically sound coastal defence alternative? A review." Aquatic Conservation: Marine and Freshwater Ecosystems 16: Telesh, I., H. Schubert en S. Skarlato (2013). "Life in the salinity gradient: Discovering mechanisms behind a new biodiversity pattern." Estuarine, Coastal and Shelf Science 135: Templeman, D., J.T. van der Wal, G. van Moorsel, M. de Kluijver, W. Lewis, A. Storm, T. van Haaren en T. Vanagt (2009). The Macrobenthic Fauna in the Dutch Sector of the North Sea in 2007 and a comparison with previous data. Amsterdam, Grontmij Aquasense. Van Dalfsen, J. en H. Holzhauer (2014a). Meetplan T0 & T1 Veldcampagne Ameland en Schiermonnikoog Delft, Deltares. Van Dalfsen, J., H. Holzhauer en E. Verduin (2014b). Veldverslag bemonstering Ameland & Schiermonnikoog 4 & 5 augustus Delft, Deltares. Van de Moortel, L. en T. Vanagt (2010). Tussentijds verslag labactiviteiten veldcampagne ecologie Ameland Stand van zaken per 1 december Oostende, ecoast. van der Spek, A. J. F., A. C. de Kruif en R. Spanhoff (2007). Richtlijnen onderwatersuppleties. Den Haag, RIKZ. van Egmond, F., W. Rooke en S. de Vries (2010). Ameland, kartering sedimentsamenstelling suppletievak en strand. Groningen, Medusa Explorations BV. Vanagt, T. en J. Heusinkveld (2010). Meetplan veldcampagne ecologie Ameland Ostende, Belgie, ecoast. Vanagt, T., L. van de Moortel, J. Heusinkveld, S. Vanden Eede, L. van Steenbrugge, G. Van Hoey en M. Vincx (2011). Veldcampagne ecologie Ameland Ostend, ecoast. Verduin, E. en L. De Vos (2014a). Analyseverslag T0 analyses Ameland & Schiermonnikoog To tbv KPP B&O Kust Ecologie. Amsterdam, Grontmij. Verduin, E. C. en L. de Vos (2014b). Analyseverslag T0 analyse Ameland & Schiermonnikoog Amsterdam, Grontmij Nederland B.V. 96
104 Verduin, E. C., L. Leewis en D. Templeman (2012). Veldcampagne Ameland Onderzoek naar de ecologische effecten van zandsuppleties op macrobenthos, epibenthos en demersale vis op Ameland, Grontmij. Verduin, E. C., L. Leewis en S. van Rooij (2011). Veldcampagne Ameland Meetplan. Amsterdam, Grontmij. Vergouwen, S. A. en H. Holzhauer (2015). Bodemdiergemeenschap Ameland en Schiermonnikoog T Delft, Deltares. Vermaas, T., E. Elias en L. Vonhögen-Peeters (2013). Ontwikkeling gefaseerde suppletie Ameland Delft, Deltares. Wijsman, J., K. Goudswaard, E. Meesters en T. Vanagt (2010). Macrofaunagemeenschap in het onderwatersuppletiegebied Ameland-midden tijdens de zomer 2009 (T0-meting), IMARES- Grontmij. 97
105
106 A Jaarlijks verschil in D50 tov 2010 Figuur A.1 Verschil in D50 tov 2010 voor de zone zeewaarts van de suppletie Figuur A.2 Verschil in D50 tov 2010 voor de suppletie zone A-1
107 Figuur A.3 Verschil in D50 tov 2010 voor de zone landwaarts van de suppletie A-2
108 B Biomassa/dichtheid per soortgroep De biomassa-dichtheid verhouding van soortgroepen van verschillende studiegebieden in de suppletiedieptezone. A B C B-1
109 D Figuur B.1 Gemiddelde biomassa (g ADG/m2)/ gemiddelde dichtheid (n/m2) per jaar in de suppletiedieptezone van phyla van de vooroever. E B-2
110 C Verspreiding Schiermonnikoog C.1 Biomassa per monsterlocatie C-1
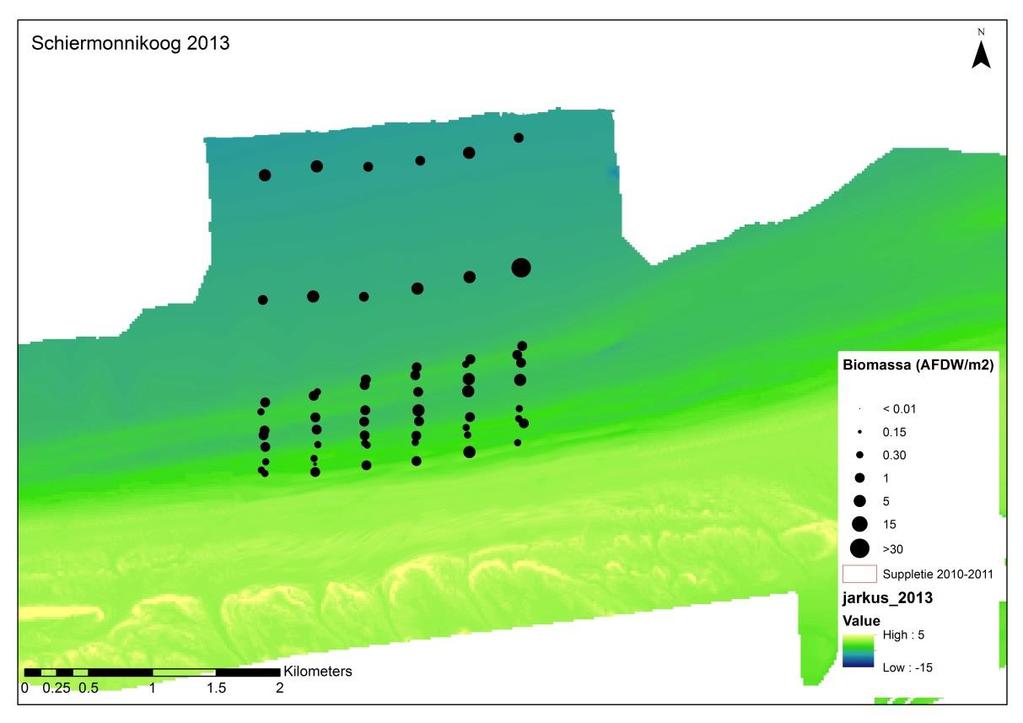
111 C-2
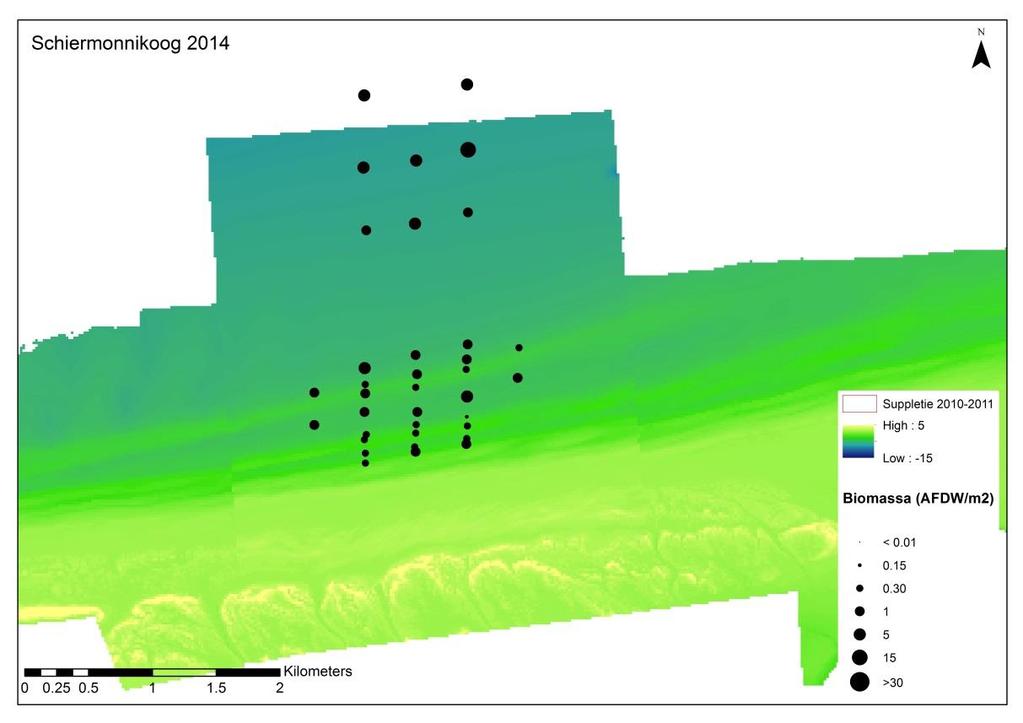
112 C-3
113 C.2 Dichtheid per monsterlocatie C-4
114 C-5
115 C-6
116 C.3 Dichtheidverspreiding van losse soorten C.3.1 Macoma balthica C-7
117 C-8
118 C.3.2 Magelona johnstoni C-9
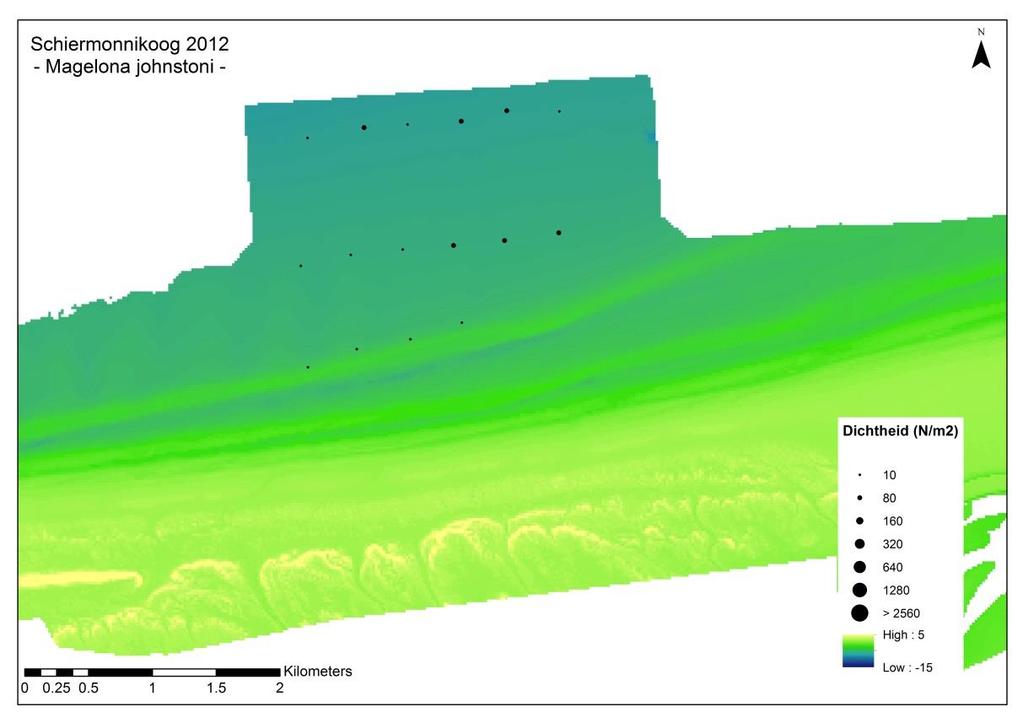
119 C-10
120 C-11
121 C.3.3 Nepthys hombergii C-12
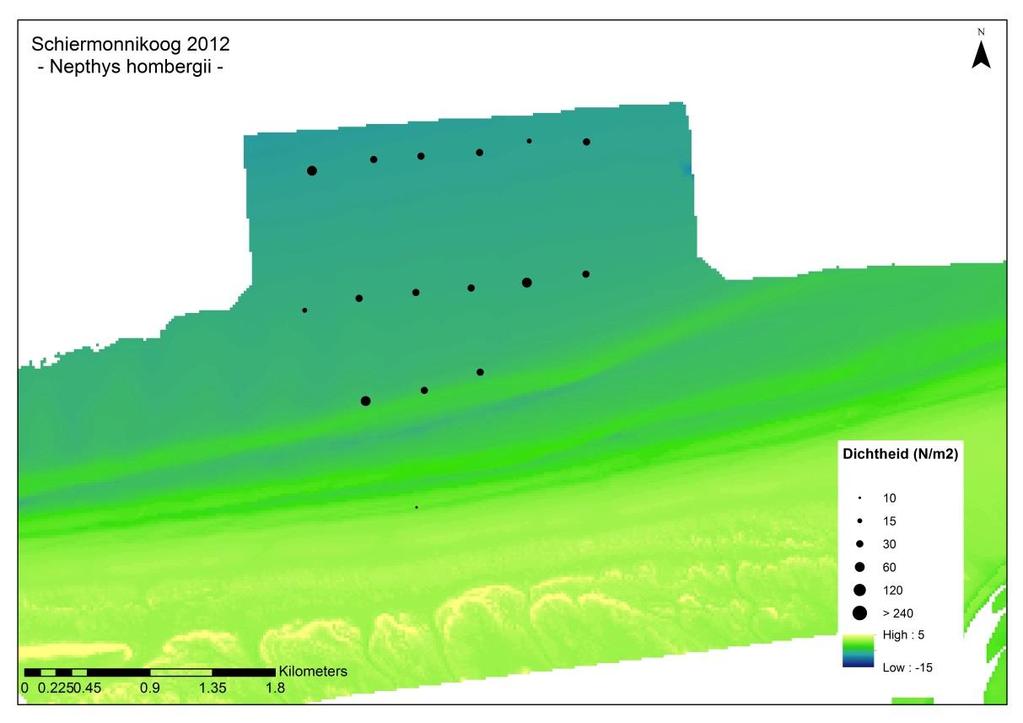
122 C-13

123 C.3.4 Echinocardium cordatum C-14
Bodemdiergemeenschap Ameland en Schiermonnikoog T0-2014
 Bodemdiergemeenschap Ameland en Schiermonnikoog T0-2014 Bodemdiergemeenschap Ameland en Schiermonnikoog T0-2014 Sophie Vergouwen Harriëtte Meijer - Holzhauer 1220040-008 Deltares, 2016, B Bodemdiergemeenschap
Bodemdiergemeenschap Ameland en Schiermonnikoog T0-2014 Bodemdiergemeenschap Ameland en Schiermonnikoog T0-2014 Sophie Vergouwen Harriëtte Meijer - Holzhauer 1220040-008 Deltares, 2016, B Bodemdiergemeenschap
Vooroever. Onderdeel van het project Ecologisch gericht suppleren
 Vooroever 1 Onderdeel van het project Ecologisch gericht suppleren Strandtent op Ameland. Foto H. Holzhauer 2014 Sinds 1990 vormen zandsuppleties een essentiële onderhoudsmaatregel voor het in stand houden
Vooroever 1 Onderdeel van het project Ecologisch gericht suppleren Strandtent op Ameland. Foto H. Holzhauer 2014 Sinds 1990 vormen zandsuppleties een essentiële onderhoudsmaatregel voor het in stand houden
Meetplan T0 & T1 Veldcampagne Ameland en Schiermonnikoog KPP B&O Kust Ecologie
 Meetplan T0 & T1 Veldcampagne Ameland en Schiermonnikoog 2014 KPP B&O Kust Ecologie Meetplan T0 & T1 Veldcampagne Ameland en Schiermonnikoog 2014 KPP B&O Kust Ecologie drs. J.A. van Dalfsen ir. H. Holzhauer
Meetplan T0 & T1 Veldcampagne Ameland en Schiermonnikoog 2014 KPP B&O Kust Ecologie Meetplan T0 & T1 Veldcampagne Ameland en Schiermonnikoog 2014 KPP B&O Kust Ecologie drs. J.A. van Dalfsen ir. H. Holzhauer
Strand. Onderdeel van het project Ecologisch gericht suppleren
 Strand 2 Onderdeel van het project Ecologisch gericht suppleren beeldbank.rws.nl / Jaap Boelens Sinds 1990 vormen zandsuppleties een essentiële onderhoudsmaatregel voor het in stand houden van onze kust.
Strand 2 Onderdeel van het project Ecologisch gericht suppleren beeldbank.rws.nl / Jaap Boelens Sinds 1990 vormen zandsuppleties een essentiële onderhoudsmaatregel voor het in stand houden van onze kust.
Kustlijnzorg in Nederland
 Kustlijnzorg in Nederland Aanleiding, uitvoering en ontwikkeling 25 september 2017 Harry de Looff RWS-WVL Inhoud Waarom Kustlijnzorg? Wat en hoe Kustlijnzorg? Ontwikkelingen en vragen 2 RWS INFORMATIE
Kustlijnzorg in Nederland Aanleiding, uitvoering en ontwikkeling 25 september 2017 Harry de Looff RWS-WVL Inhoud Waarom Kustlijnzorg? Wat en hoe Kustlijnzorg? Ontwikkelingen en vragen 2 RWS INFORMATIE
Analyseverslag T0 analyse Ameland & Schiermonnikoog
 Analyseverslag T0 analyse Ameland & Schiermonnikoog 2014-8 T0, t.b.v. KPP B&O Kust Ecologie Deltares Grontmij Nederland B.V. Amsterdam, 25 september 2014 Verantwoording Titel : Analyseverslag T0 analyse
Analyseverslag T0 analyse Ameland & Schiermonnikoog 2014-8 T0, t.b.v. KPP B&O Kust Ecologie Deltares Grontmij Nederland B.V. Amsterdam, 25 september 2014 Verantwoording Titel : Analyseverslag T0 analyse
Ondiepe Kust, Schiermonnikoog, Egmond, Macrobenthos, Medusa, 2005
 Laan van Westenenk 501 Postbus 342 7300 AH Apeldoorn The Netherlands TNO- B&O-DH Zandige kust www.tno.nl T +31 55 549 34 93 F +31 55 549 39 01 info@mep.tno.nl Date 18 th Oktober 2005 Author J.A. van Dalfsen
Laan van Westenenk 501 Postbus 342 7300 AH Apeldoorn The Netherlands TNO- B&O-DH Zandige kust www.tno.nl T +31 55 549 34 93 F +31 55 549 39 01 info@mep.tno.nl Date 18 th Oktober 2005 Author J.A. van Dalfsen
Lichamelijke factoren als voorspeller voor psychisch. en lichamelijk herstel bij anorexia nervosa. Physical factors as predictors of psychological and
 Lichamelijke factoren als voorspeller voor psychisch en lichamelijk herstel bij anorexia nervosa Physical factors as predictors of psychological and physical recovery of anorexia nervosa Liesbeth Libbers
Lichamelijke factoren als voorspeller voor psychisch en lichamelijk herstel bij anorexia nervosa Physical factors as predictors of psychological and physical recovery of anorexia nervosa Liesbeth Libbers
Stand van zaken onderzoeken medio 2018
 Stand van zaken onderzoeken medio 2018 Over Natuurlijk Veilig Natuurlijk Veilig is een meerjarig samenwerkingsproject van Rijkswaterstaat, de Wadden vereniging, Staatsbosbeheer, Vogelbescherming Nederland,
Stand van zaken onderzoeken medio 2018 Over Natuurlijk Veilig Natuurlijk Veilig is een meerjarig samenwerkingsproject van Rijkswaterstaat, de Wadden vereniging, Staatsbosbeheer, Vogelbescherming Nederland,
Ecologische effecten suppletie Ameland Interim rapportage ihkv KPP B&O Kust Ecologie
 Ecologische effecten suppletie Ameland 2009-2012 Interim rapportage ihkv KPP B&O Kust Ecologie Ecologische effecten suppletie Ameland 2009-2012 Interim rapportage ihkv KPP B&O Kust Ecologie ir. H. Holzhauer
Ecologische effecten suppletie Ameland 2009-2012 Interim rapportage ihkv KPP B&O Kust Ecologie Ecologische effecten suppletie Ameland 2009-2012 Interim rapportage ihkv KPP B&O Kust Ecologie ir. H. Holzhauer
Kenmerk ZKS Doorkiesnummer +31 (0) (0)
 (0)") Memo Aan Jasper Fiselier (DHV) Datum Van Pieter Koen Tonnon Jan Mulder Kenmerk Doorkiesnummer +31 (0)88 33 58 464 +31 (0)88 33 58 446 Aantal pagina's 9 E-mail pieterkoen.tonnon @deltares.nl jan.mulder@deltares.nl
Memo Aan Jasper Fiselier (DHV) Datum Van Pieter Koen Tonnon Jan Mulder Kenmerk Doorkiesnummer +31 (0)88 33 58 464 +31 (0)88 33 58 446 Aantal pagina's 9 E-mail pieterkoen.tonnon @deltares.nl jan.mulder@deltares.nl
CHROMA STANDAARDREEKS
 CHROMA STANDAARDREEKS Chroma-onderzoeken Een chroma geeft een beeld over de kwaliteit van bijvoorbeeld een bodem of compost. Een chroma bestaat uit 4 zones. Uit elke zone is een bepaald kwaliteitsaspect
CHROMA STANDAARDREEKS Chroma-onderzoeken Een chroma geeft een beeld over de kwaliteit van bijvoorbeeld een bodem of compost. Een chroma bestaat uit 4 zones. Uit elke zone is een bepaald kwaliteitsaspect
De tor. Memo. Aan Jaap Starke (Rijkswaterstaat) Datum 5 februari Kenmerk ZKS Aantal pagina's 10. Van Reinier Schrijvershof
 Datum 5 februari Kenmerk ZKS Aantal pagina's 10. Van Reinier Schrijvershof") Memo De tor s Aan Jaap Starke (Rijkswaterstaat) Datum Van Reinier Schrijvershof Kenmerk Doorkiesnummer +31(0)88335 7483 Aantal pagina's 10 E-mail reinier.schrijvershof@deltares.nl Onderwerp Verplaatsing
Memo De tor s Aan Jaap Starke (Rijkswaterstaat) Datum Van Reinier Schrijvershof Kenmerk Doorkiesnummer +31(0)88335 7483 Aantal pagina's 10 E-mail reinier.schrijvershof@deltares.nl Onderwerp Verplaatsing
De Relatie tussen de Fysieke Omgeving en het Beweeggedrag van Kinderen gebruik. makend van GPS- en Versnellingsmeterdata
 De Relatie tussen de Fysieke Omgeving en het Beweeggedrag van Kinderen gebruik makend van GPS- en Versnellingsmeterdata The relationship Between the Physical Environment and Physical Activity in Children
De Relatie tussen de Fysieke Omgeving en het Beweeggedrag van Kinderen gebruik makend van GPS- en Versnellingsmeterdata The relationship Between the Physical Environment and Physical Activity in Children
De causale Relatie tussen Intimiteit en Seksueel verlangen en de. modererende invloed van Sekse en Relatietevredenheid op deze relatie
 Causale Relatie tussen intimiteit en seksueel verlangen 1 De causale Relatie tussen Intimiteit en Seksueel verlangen en de modererende invloed van Sekse en Relatietevredenheid op deze relatie The causal
Causale Relatie tussen intimiteit en seksueel verlangen 1 De causale Relatie tussen Intimiteit en Seksueel verlangen en de modererende invloed van Sekse en Relatietevredenheid op deze relatie The causal
COGNITIEVE DISSONANTIE EN ROKERS COGNITIVE DISSONANCE AND SMOKERS
 COGNITIEVE DISSONANTIE EN ROKERS Gezondheidsgedrag als compensatie voor de schadelijke gevolgen van roken COGNITIVE DISSONANCE AND SMOKERS Health behaviour as compensation for the harmful effects of smoking
COGNITIEVE DISSONANTIE EN ROKERS Gezondheidsgedrag als compensatie voor de schadelijke gevolgen van roken COGNITIVE DISSONANCE AND SMOKERS Health behaviour as compensation for the harmful effects of smoking
Ecologisch gericht suppleren I
 Ecologisch gericht suppleren I resultaten van het onderzoek Petra Damsma Harriette Holzhauer Tommer Vermaas Bert vd Valk Luca van Duren Annelies de Backer inhoud Het kustbeleid en kustbeheer door RWS De
Ecologisch gericht suppleren I resultaten van het onderzoek Petra Damsma Harriette Holzhauer Tommer Vermaas Bert vd Valk Luca van Duren Annelies de Backer inhoud Het kustbeleid en kustbeheer door RWS De
Memo. 1 Introductie. +31(0) Kees den Heijer Roeland de Zeeuw (Shore Monitoring & Research)
 Kees den Heijer Roeland de Zeeuw (Shore Monitoring & Research)") Memo Aan Saskia Huijs (Rijkswaterstaat Zee en Delta) Datum Kenmerk Doorkiesnummer Van +31(0)88335 7945 Kees den Heijer Roeland de Zeeuw (Shore Monitoring & Research) Aantal pagina's 9 E-mail kees.denheijer
Memo Aan Saskia Huijs (Rijkswaterstaat Zee en Delta) Datum Kenmerk Doorkiesnummer Van +31(0)88335 7945 Kees den Heijer Roeland de Zeeuw (Shore Monitoring & Research) Aantal pagina's 9 E-mail kees.denheijer
van Werknemers Well-being Drs. P.E. Gouw
 De Invloed van Werk- en Persoonskenmerken op het Welbevinden van Werknemers The Influence of Job and Personality Characteristics on Employee Well-being Drs. P.E. Gouw Eerste begeleider: Dr. S. van Hooren
De Invloed van Werk- en Persoonskenmerken op het Welbevinden van Werknemers The Influence of Job and Personality Characteristics on Employee Well-being Drs. P.E. Gouw Eerste begeleider: Dr. S. van Hooren
VELDVERSLAG AMELAND EN SCHIERMONNIKOOG 2013
 VELDVERSLAG AMELAND EN SCHIERMONNIKOOG 2013 BEMONSTERING (JUVENIELE) DEMERSALE VIS ILVO MEDEDELING nr 140 oktober 2013 Annelies De Backer Gert Van Hoey Veldverslag Ameland en Schiermonnikoog 2013 Bemonstering
VELDVERSLAG AMELAND EN SCHIERMONNIKOOG 2013 BEMONSTERING (JUVENIELE) DEMERSALE VIS ILVO MEDEDELING nr 140 oktober 2013 Annelies De Backer Gert Van Hoey Veldverslag Ameland en Schiermonnikoog 2013 Bemonstering
Het Effect van Verschil in Sociale Invloed van Ouders en Vrienden op het Alcoholgebruik van Adolescenten.
 Het Effect van Verschil in Sociale Invloed van Ouders en Vrienden op het Alcoholgebruik van Adolescenten. The Effect of Difference in Peer and Parent Social Influences on Adolescent Alcohol Use. Nadine
Het Effect van Verschil in Sociale Invloed van Ouders en Vrienden op het Alcoholgebruik van Adolescenten. The Effect of Difference in Peer and Parent Social Influences on Adolescent Alcohol Use. Nadine
Meetplan veldcampagne ecologie Ameland 2010
 Meetplan veldcampagne ecologie Ameland 2010 Thomas Vanagt en Jannes Heusinkveld 01/11/2010 eindversie Contents 1. Inleiding... 3 1.1 Achtergrond onderzoeksprogramma effecten suppleties... 3 1.2 Veldcampgane
Meetplan veldcampagne ecologie Ameland 2010 Thomas Vanagt en Jannes Heusinkveld 01/11/2010 eindversie Contents 1. Inleiding... 3 1.1 Achtergrond onderzoeksprogramma effecten suppleties... 3 1.2 Veldcampgane
Outline A PERMANENT PASTURE LAYER BASED ON OPEN DATA 11/24/2014. The creation and monitoring of a permanent pasture layer
 A PERMANENT PASTURE LAYER BASED ON OPEN DATA The creation and monitoring of a permanent pasture layer 20 th of November 2014, Marcel Meijer Outline Open Data in the Netherland Greening elements Calculating
A PERMANENT PASTURE LAYER BASED ON OPEN DATA The creation and monitoring of a permanent pasture layer 20 th of November 2014, Marcel Meijer Outline Open Data in the Netherland Greening elements Calculating
MyDHL+ Van Non-Corporate naar Corporate
 MyDHL+ Van Non-Corporate naar Corporate Van Non-Corporate naar Corporate In MyDHL+ is het mogelijk om meerdere gebruikers aan uw set-up toe te voegen. Wanneer er bijvoorbeeld meerdere collega s van dezelfde
MyDHL+ Van Non-Corporate naar Corporate Van Non-Corporate naar Corporate In MyDHL+ is het mogelijk om meerdere gebruikers aan uw set-up toe te voegen. Wanneer er bijvoorbeeld meerdere collega s van dezelfde
De Relatie tussen Betrokkenheid bij Pesten en Welbevinden en de Invloed van Sociale Steun en. Discrepantie
 De Relatie tussen Betrokkenheid bij Pesten en Welbevinden en de Invloed van Sociale Steun en Discrepantie The Relationship between Involvement in Bullying and Well-Being and the Influence of Social Support
De Relatie tussen Betrokkenheid bij Pesten en Welbevinden en de Invloed van Sociale Steun en Discrepantie The Relationship between Involvement in Bullying and Well-Being and the Influence of Social Support
EVALUATIE VAN DE VOOROEVERSUPPLETIE BIJ HEEMSKERK (2011-2012) EINDVERSLAG, BACHELOR EINDOPDRACHT, CIVIELE TECHNIEK, UNIVERSITEIT TWENTE
 EINDVERSLAG, BACHELOR EINDOPDRACHT, CIVIELE TECHNIEK, UNIVERSITEIT TWENTE") EVALUATIE VAN DE VOOROEVERSUPPLETIE BIJ HEEMSKERK (2011-2012) EINDVERSLAG, BACHELOR EINDOPDRACHT, CIVIELE TECHNIEK, UNIVERSITEIT TWENTE Daan Boot April 2015 Juni 2015 Evaluatie van de vooroeversuppletie
EVALUATIE VAN DE VOOROEVERSUPPLETIE BIJ HEEMSKERK (2011-2012) EINDVERSLAG, BACHELOR EINDOPDRACHT, CIVIELE TECHNIEK, UNIVERSITEIT TWENTE Daan Boot April 2015 Juni 2015 Evaluatie van de vooroeversuppletie
De Invloed van Dagelijkse Stress op Burn-Out Klachten, Gemodereerd door Mentale. Veerkracht en Demografische Variabelen
 Running head: INVLOED VAN DAGELIJKSE STRESS OP BURN-OUT KLACHTEN De Invloed van Dagelijkse Stress op Burn-Out Klachten, Gemodereerd door Mentale Veerkracht en Demografische Variabelen The Influence of
Running head: INVLOED VAN DAGELIJKSE STRESS OP BURN-OUT KLACHTEN De Invloed van Dagelijkse Stress op Burn-Out Klachten, Gemodereerd door Mentale Veerkracht en Demografische Variabelen The Influence of
Basiskustlijn Herziening van de ligging van de basiskustlijn. Datum 30 januari 2018
 Basiskustlijn 2017 Herziening van de ligging van de basiskustlijn Datum 30 januari 2018 Status Definitief 1 Colofon Bestuurskern Dir. Algemeen Waterbeleid en Veiligheid Den Haag Contactpersoon F.P. Hallie
Basiskustlijn 2017 Herziening van de ligging van de basiskustlijn Datum 30 januari 2018 Status Definitief 1 Colofon Bestuurskern Dir. Algemeen Waterbeleid en Veiligheid Den Haag Contactpersoon F.P. Hallie
Sekseverschillen in Huilfrequentie en Psychosociale Problemen. bij Schoolgaande Kinderen van 6 tot 10 jaar
 Sekseverschillen in Huilfrequentie en Psychosociale Problemen bij Schoolgaande Kinderen van 6 tot 10 jaar Gender Differences in Crying Frequency and Psychosocial Problems in Schoolgoing Children aged 6
Sekseverschillen in Huilfrequentie en Psychosociale Problemen bij Schoolgaande Kinderen van 6 tot 10 jaar Gender Differences in Crying Frequency and Psychosocial Problems in Schoolgoing Children aged 6
De Relatie Tussen de Gehanteerde Copingstijl en Pesten op het Werk. The Relation Between the Used Coping Style and Bullying at Work.
 De Relatie Tussen de Gehanteerde Copingstijl en Pesten op het Werk The Relation Between the Used Coping Style and Bullying at Work Merijn Daerden Studentnummer: 850225144 Werkstuk: Empirisch afstudeeronderzoek:
De Relatie Tussen de Gehanteerde Copingstijl en Pesten op het Werk The Relation Between the Used Coping Style and Bullying at Work Merijn Daerden Studentnummer: 850225144 Werkstuk: Empirisch afstudeeronderzoek:
Invloed van het aantal kinderen op de seksdrive en relatievoorkeur
 Invloed van het aantal kinderen op de seksdrive en relatievoorkeur M. Zander MSc. Eerste begeleider: Tweede begeleider: dr. W. Waterink drs. J. Eshuis Oktober 2014 Faculteit Psychologie en Onderwijswetenschappen
Invloed van het aantal kinderen op de seksdrive en relatievoorkeur M. Zander MSc. Eerste begeleider: Tweede begeleider: dr. W. Waterink drs. J. Eshuis Oktober 2014 Faculteit Psychologie en Onderwijswetenschappen
Pesten onder Leerlingen met Autisme Spectrum Stoornissen op de Middelbare School: de Participantrollen en het Verband met de Theory of Mind.
 Pesten onder Leerlingen met Autisme Spectrum Stoornissen op de Middelbare School: de Participantrollen en het Verband met de Theory of Mind. Bullying among Students with Autism Spectrum Disorders in Secondary
Pesten onder Leerlingen met Autisme Spectrum Stoornissen op de Middelbare School: de Participantrollen en het Verband met de Theory of Mind. Bullying among Students with Autism Spectrum Disorders in Secondary
Het executief en het sociaal cognitief functioneren bij licht verstandelijk. gehandicapte jeugdigen. Samenhang met emotionele- en gedragsproblemen
 Het executief en het sociaal cognitief functioneren bij licht verstandelijk gehandicapte jeugdigen. Samenhang met emotionele- en gedragsproblemen Executive and social cognitive functioning of mentally
Het executief en het sociaal cognitief functioneren bij licht verstandelijk gehandicapte jeugdigen. Samenhang met emotionele- en gedragsproblemen Executive and social cognitive functioning of mentally
Introductie in flowcharts
 Introductie in flowcharts Flow Charts Een flow chart kan gebruikt worden om: Processen definieren en analyseren. Een beeld vormen van een proces voor analyse, discussie of communicatie. Het definieren,
Introductie in flowcharts Flow Charts Een flow chart kan gebruikt worden om: Processen definieren en analyseren. Een beeld vormen van een proces voor analyse, discussie of communicatie. Het definieren,
Effecten van contactgericht spelen en leren op de ouder-kindrelatie bij autisme
 Effecten van contactgericht spelen en leren op de ouder-kindrelatie bij autisme Effects of Contact-oriented Play and Learning in the Relationship between parent and child with autism Kristel Stes Studentnummer:
Effecten van contactgericht spelen en leren op de ouder-kindrelatie bij autisme Effects of Contact-oriented Play and Learning in the Relationship between parent and child with autism Kristel Stes Studentnummer:
Beïnvloedt Gentle Teaching Vaardigheden van Begeleiders en Companionship en Angst bij Verstandelijk Beperkte Cliënten?
 Beïnvloedt Gentle Teaching Vaardigheden van Begeleiders en Companionship en Angst bij Verstandelijk Beperkte Cliënten? Does Gentle Teaching have Effect on Skills of Caregivers and Companionship and Anxiety
Beïnvloedt Gentle Teaching Vaardigheden van Begeleiders en Companionship en Angst bij Verstandelijk Beperkte Cliënten? Does Gentle Teaching have Effect on Skills of Caregivers and Companionship and Anxiety
Meten om te weten: 2,5 jaar Zandmotor
 : 2,5 jaar Carola van Gelder-Maas Projectmanager WVL Rijkswaterstaat 31 maart 2014 Hoe zat het ook alweer? Eroderende kustlijn NL kust 12 Mm³ zandsuppleties per jaar Zeespiegelstijging Zwakke schakels
: 2,5 jaar Carola van Gelder-Maas Projectmanager WVL Rijkswaterstaat 31 maart 2014 Hoe zat het ook alweer? Eroderende kustlijn NL kust 12 Mm³ zandsuppleties per jaar Zeespiegelstijging Zwakke schakels
Mentaal Weerbaar Blauw
 Mentaal Weerbaar Blauw de invloed van stereotypen over etnische minderheden cynisme en negatieve emoties op de mentale weerbaarheid van politieagenten begeleiders: dr. Anita Eerland & dr. Arjan Bos dr.
Mentaal Weerbaar Blauw de invloed van stereotypen over etnische minderheden cynisme en negatieve emoties op de mentale weerbaarheid van politieagenten begeleiders: dr. Anita Eerland & dr. Arjan Bos dr.
bij Kinderen met een Ernstige Vorm van Dyslexie of Children with a Severe Form of Dyslexia Ans van Velthoven
 Neuropsychologische Behandeling en Sociaal Emotioneel Welzijn bij Kinderen met een Ernstige Vorm van Dyslexie Neuropsychological Treatment and Social Emotional Well-being of Children with a Severe Form
Neuropsychologische Behandeling en Sociaal Emotioneel Welzijn bij Kinderen met een Ernstige Vorm van Dyslexie Neuropsychological Treatment and Social Emotional Well-being of Children with a Severe Form
FOR DUTCH STUDENTS! ENGLISH VERSION NEXT PAGE. Toets Inleiding Kansrekening 1 8 februari 2010
 FOR DUTCH STUDENTS! ENGLISH VERSION NEXT PAGE Toets Inleiding Kansrekening 1 8 februari 2010 Voeg aan het antwoord van een opgave altijd het bewijs, de berekening of de argumentatie toe. Als je een onderdeel
FOR DUTCH STUDENTS! ENGLISH VERSION NEXT PAGE Toets Inleiding Kansrekening 1 8 februari 2010 Voeg aan het antwoord van een opgave altijd het bewijs, de berekening of de argumentatie toe. Als je een onderdeel
Bijlage 2: Onderbouwing concept suppletieprogramma 2016-2019
 Bijlage 2: Onderbouwing concept suppletieprogramma 2016-2019 1 Concept suppletieprogramma 2016-2019 n.a.v. toetsing basiskustlijn Uit de toetsing van de kustlijn blijkt dat de basiskustlijn (BKL) tussen
Bijlage 2: Onderbouwing concept suppletieprogramma 2016-2019 1 Concept suppletieprogramma 2016-2019 n.a.v. toetsing basiskustlijn Uit de toetsing van de kustlijn blijkt dat de basiskustlijn (BKL) tussen
GOVERNMENT NOTICE. STAATSKOERANT, 18 AUGUSTUS 2017 No NATIONAL TREASURY. National Treasury/ Nasionale Tesourie NO AUGUST
 National Treasury/ Nasionale Tesourie 838 Local Government: Municipal Finance Management Act (56/2003): Draft Amendments to Municipal Regulations on Minimum Competency Levels, 2017 41047 GOVERNMENT NOTICE
National Treasury/ Nasionale Tesourie 838 Local Government: Municipal Finance Management Act (56/2003): Draft Amendments to Municipal Regulations on Minimum Competency Levels, 2017 41047 GOVERNMENT NOTICE
De Invloed van Perceived Severity op Condoomgebruik en HIV-Testgedrag. The Influence of Perceived Severity on Condom Use and HIV-Testing Behavior
 De Invloed van Perceived Severity op Condoomgebruik en HIV-Testgedrag The Influence of Perceived Severity on Condom Use and HIV-Testing Behavior Martin. W. van Duijn Student: 838797266 Eerste begeleider:
De Invloed van Perceived Severity op Condoomgebruik en HIV-Testgedrag The Influence of Perceived Severity on Condom Use and HIV-Testing Behavior Martin. W. van Duijn Student: 838797266 Eerste begeleider:
Ecobeach. Een duurzaam strand door drainage. Brouwersdam, 3-10-2014. Bas Reedijk. Hoofd afdeling kustwaterbouw BAM Infraconsult
 Ecobeach Een duurzaam strand door drainage Brouwersdam, 3-10-2014 Bas Reedijk Hoofd afdeling kustwaterbouw BAM Infraconsult Inhoud Wat is Ecobeach De ervaring met Ecobeach bij Egmond Toepassing van Ecobeach
Ecobeach Een duurzaam strand door drainage Brouwersdam, 3-10-2014 Bas Reedijk Hoofd afdeling kustwaterbouw BAM Infraconsult Inhoud Wat is Ecobeach De ervaring met Ecobeach bij Egmond Toepassing van Ecobeach
KLZ-eco en het MLT Programma: ecologisch gericht suppleren Bert van der Valk en Harriette Holzhauer
 KLZ-eco en het MLT Programma: ecologisch gericht suppleren 2009-2015 Bert van der Valk en Harriette Holzhauer Vanuit het recente verleden KLZ 2008: Review van effecten van gangbare suppletie activiteit
KLZ-eco en het MLT Programma: ecologisch gericht suppleren 2009-2015 Bert van der Valk en Harriette Holzhauer Vanuit het recente verleden KLZ 2008: Review van effecten van gangbare suppletie activiteit
DANKBAARHEID, PSYCHOLOGISCHE BASISBEHOEFTEN EN LEVENSDOELEN 1
 DANKBAARHEID, PSYCHOLOGISCHE BASISBEHOEFTEN EN LEVENSDOELEN 1 Dankbaarheid in Relatie tot Intrinsieke Levensdoelen: Het mediërende Effect van Psychologische Basisbehoeften Karin Nijssen Open Universiteit
DANKBAARHEID, PSYCHOLOGISCHE BASISBEHOEFTEN EN LEVENSDOELEN 1 Dankbaarheid in Relatie tot Intrinsieke Levensdoelen: Het mediërende Effect van Psychologische Basisbehoeften Karin Nijssen Open Universiteit
Zandsuppleties en Morfologie langs de Nederlandse kust
 Zandsuppleties en Morfologie langs de Nederlandse kust Sander Boer, Landelijke Kustdagen 2014 Inhoud presentatie Verankering in beleid en wetgeving Kustlijnzorg Toekomstige uitdaging Kustversterking Hondsbossche
Zandsuppleties en Morfologie langs de Nederlandse kust Sander Boer, Landelijke Kustdagen 2014 Inhoud presentatie Verankering in beleid en wetgeving Kustlijnzorg Toekomstige uitdaging Kustversterking Hondsbossche
Wat is de Modererende Rol van Consciëntieusheid, Extraversie en Neuroticisme op de Relatie tussen Depressieve Symptomen en Overeten?
 De Modererende rol van Persoonlijkheid op de Relatie tussen Depressieve Symptomen en Overeten 1 Wat is de Modererende Rol van Consciëntieusheid, Extraversie en Neuroticisme op de Relatie tussen Depressieve
De Modererende rol van Persoonlijkheid op de Relatie tussen Depressieve Symptomen en Overeten 1 Wat is de Modererende Rol van Consciëntieusheid, Extraversie en Neuroticisme op de Relatie tussen Depressieve
(1) De hoofdfunctie van ons gezelschap is het aanbieden van onderwijs. (2) Ons gezelschap is er om kunsteducatie te verbeteren
 De hoofdfunctie van ons gezelschap is het aanbieden van onderwijs. (2) Ons gezelschap is er om kunsteducatie te verbeteren") (1) De hoofdfunctie van ons gezelschap is het aanbieden van onderwijs (2) Ons gezelschap is er om kunsteducatie te verbeteren (3) Ons gezelschap helpt gemeenschappen te vormen en te binden (4) De producties
(1) De hoofdfunctie van ons gezelschap is het aanbieden van onderwijs (2) Ons gezelschap is er om kunsteducatie te verbeteren (3) Ons gezelschap helpt gemeenschappen te vormen en te binden (4) De producties
FOR DUTCH STUDENTS! ENGLISH VERSION NEXT PAGE
 FOR DUTCH STUDENTS! ENGLISH VERSION NEXT PAGE Tentamen Bewijzen en Technieken 1 7 januari 211, duur 3 uur. Voeg aan het antwoord van een opgave altijd het bewijs, de berekening of de argumentatie toe.
FOR DUTCH STUDENTS! ENGLISH VERSION NEXT PAGE Tentamen Bewijzen en Technieken 1 7 januari 211, duur 3 uur. Voeg aan het antwoord van een opgave altijd het bewijs, de berekening of de argumentatie toe.
Korrelgrootte karakteristiek van het strand
 Korrelgrootte karakteristiek van het strand Effecten van suppleties op het sediment en profiel van het strand Afstudeerscriptie Ruben de Vries augustus 2009 Korrelgrootte karakteristiek van het strand
Korrelgrootte karakteristiek van het strand Effecten van suppleties op het sediment en profiel van het strand Afstudeerscriptie Ruben de Vries augustus 2009 Korrelgrootte karakteristiek van het strand
De Relatie tussen Lichamelijke Gezondheid, Veerkracht en Subjectief. Welbevinden bij Inwoners van Serviceflats
 De Relatie tussen Lichamelijke Gezondheid, Veerkracht en Subjectief Welbevinden bij Inwoners van Serviceflats The Relationship between Physical Health, Resilience and Subjective Wellbeing of Inhabitants
De Relatie tussen Lichamelijke Gezondheid, Veerkracht en Subjectief Welbevinden bij Inwoners van Serviceflats The Relationship between Physical Health, Resilience and Subjective Wellbeing of Inhabitants
The relationship between social support and loneliness and depressive symptoms in Turkish elderly: the mediating role of the ability to cope
 The relationship between social support and loneliness and depressive symptoms in Turkish elderly: the mediating role of the ability to cope Een onderzoek naar de relatie tussen sociale steun en depressieve-
The relationship between social support and loneliness and depressive symptoms in Turkish elderly: the mediating role of the ability to cope Een onderzoek naar de relatie tussen sociale steun en depressieve-
Summary 124
 Summary Summary 124 Summary Summary Corporate social responsibility and current legislation encourage the employment of people with disabilities in inclusive organizations. However, people with disabilities
Summary Summary 124 Summary Summary Corporate social responsibility and current legislation encourage the employment of people with disabilities in inclusive organizations. However, people with disabilities
Relatie tussen Persoonlijkheid, Opleidingsniveau, Leeftijd, Geslacht en Korte- en Lange- Termijn Seksuele Strategieën
 Relatie tussen Persoonlijkheid, Opleidingsniveau, Leeftijd, Geslacht en Korte- en Lange- Termijn Seksuele Strategieën The Relation between Personality, Education, Age, Sex and Short- and Long- Term Sexual
Relatie tussen Persoonlijkheid, Opleidingsniveau, Leeftijd, Geslacht en Korte- en Lange- Termijn Seksuele Strategieën The Relation between Personality, Education, Age, Sex and Short- and Long- Term Sexual
Werk in balans. verloop bij verzorgenden en verpleegkundigen. Work in balance. turnover of nurses and health-care workers.
 Werk in balans Een onderzoek naar de invloed van werktijden op werkthuisinterferentie en de gevolgen daarvan voor burnout en verloop bij verzorgenden en verpleegkundigen. Work in balance A study of the
Werk in balans Een onderzoek naar de invloed van werktijden op werkthuisinterferentie en de gevolgen daarvan voor burnout en verloop bij verzorgenden en verpleegkundigen. Work in balance A study of the
De Relatie Tussen Persoonskenmerken en Ervaren Lijden bij. Verslaafde Patiënten met PTSS
 Persoonskenmerken en ervaren lijden bij verslaving en PTSS 1 De Relatie Tussen Persoonskenmerken en Ervaren Lijden bij Verslaafde Patiënten met PTSS The Relationship between Personality Traits and Suffering
Persoonskenmerken en ervaren lijden bij verslaving en PTSS 1 De Relatie Tussen Persoonskenmerken en Ervaren Lijden bij Verslaafde Patiënten met PTSS The Relationship between Personality Traits and Suffering
OVERGANGSREGELS / TRANSITION RULES 2007/2008
 OVERGANGSREGELS / TRANSITION RULES 2007/2008 Instructie Met als doel het studiecurriculum te verbeteren of verduidelijken heeft de faculteit FEB besloten tot aanpassingen in enkele programma s die nu van
OVERGANGSREGELS / TRANSITION RULES 2007/2008 Instructie Met als doel het studiecurriculum te verbeteren of verduidelijken heeft de faculteit FEB besloten tot aanpassingen in enkele programma s die nu van
een andere kijk op klimaatadaptatie
 een andere kijk op klimaatadaptatie Jan Mulder datum 24 mei 2011 Klimaatverandering: WATER perspectief dominant Observed changes "Warming of the climate system is unequivocal, as is now evident from observations
een andere kijk op klimaatadaptatie Jan Mulder datum 24 mei 2011 Klimaatverandering: WATER perspectief dominant Observed changes "Warming of the climate system is unequivocal, as is now evident from observations
Morfologische ontwikkeling van de kust van Voorne. Quirijn Lodder
 Morfologische ontwikkeling van de kust van Voorne Quirijn Lodder Inhoudsopgave 1. Kustontwikkeling Nederland 2. Kustontwikkeling Zuidwestelijke Delta 3. Kustlijnzorg 4. Samenvatting 1. Kustontwikkeling
Morfologische ontwikkeling van de kust van Voorne Quirijn Lodder Inhoudsopgave 1. Kustontwikkeling Nederland 2. Kustontwikkeling Zuidwestelijke Delta 3. Kustlijnzorg 4. Samenvatting 1. Kustontwikkeling
Het Verband Tussen Persoonlijkheid, Stress en Coping. The Relation Between Personality, Stress and Coping
 Het Verband Tussen Persoonlijkheid, Stress en Coping The Relation Between Personality, Stress and Coping J.R.M. de Vos Oktober 2009 1e begeleider: Mw. Dr. T. Houtmans 2e begeleider: Mw. Dr. K. Proost Faculteit
Het Verband Tussen Persoonlijkheid, Stress en Coping The Relation Between Personality, Stress and Coping J.R.M. de Vos Oktober 2009 1e begeleider: Mw. Dr. T. Houtmans 2e begeleider: Mw. Dr. K. Proost Faculteit
Adherence aan HWO en meer bewegen
 Adherence aan HWO en meer bewegen Een experimenteel onderzoek naar de effecten van het motivationele stadium van patiënten en de adherence aan huiswerkoefeningen (HWO) bij fysiotherapie en het meer bewegen.
Adherence aan HWO en meer bewegen Een experimenteel onderzoek naar de effecten van het motivationele stadium van patiënten en de adherence aan huiswerkoefeningen (HWO) bij fysiotherapie en het meer bewegen.
TECHNISCHE UNIVERSITEIT EINDHOVEN Faculteit Wiskunde en Informatica. Examination 2DL04 Friday 16 november 2007, hours.
 TECHNISCHE UNIVERSITEIT EINDHOVEN Faculteit Wiskunde en Informatica Examination 2DL04 Friday 16 november 2007, 14.00-17.00 hours. De uitwerkingen van de opgaven dienen duidelijk geformuleerd en overzichtelijk
TECHNISCHE UNIVERSITEIT EINDHOVEN Faculteit Wiskunde en Informatica Examination 2DL04 Friday 16 november 2007, 14.00-17.00 hours. De uitwerkingen van de opgaven dienen duidelijk geformuleerd en overzichtelijk
Climate impact on fish productivity: key mechanisms in North Sea plaice
 Climate impact on fish productivity: key mechanisms in North Sea plaice A.D. Rijnsdorp, R. van Hal, M. Hufnagl, R.D.M. Nash, A. Schroeder, L.R. Teal, I. Tulp, R. Witbaard, D. Beare, H.W. van der Veer Wageningen
Climate impact on fish productivity: key mechanisms in North Sea plaice A.D. Rijnsdorp, R. van Hal, M. Hufnagl, R.D.M. Nash, A. Schroeder, L.R. Teal, I. Tulp, R. Witbaard, D. Beare, H.W. van der Veer Wageningen
General info on using shopping carts with Ingenico epayments
 Inhoudsopgave 1. Disclaimer 2. What is a PSPID? 3. What is an API user? How is it different from other users? 4. What is an operation code? And should I choose "Authorisation" or "Sale"? 5. What is an
Inhoudsopgave 1. Disclaimer 2. What is a PSPID? 3. What is an API user? How is it different from other users? 4. What is an operation code? And should I choose "Authorisation" or "Sale"? 5. What is an
Impact en disseminatie. Saskia Verhagen Franka vd Wijdeven
 Impact en disseminatie Saskia Verhagen Franka vd Wijdeven Wie is wie? Voorstel rondje Wat hoop je te leren? Heb je iets te delen? Wat zegt de Programma Gids? WHAT DO IMPACT AND SUSTAINABILITY MEAN? Impact
Impact en disseminatie Saskia Verhagen Franka vd Wijdeven Wie is wie? Voorstel rondje Wat hoop je te leren? Heb je iets te delen? Wat zegt de Programma Gids? WHAT DO IMPACT AND SUSTAINABILITY MEAN? Impact
Het Verband Tussen Negatieve Levensgebeurtenissen, 5-HTTLPR en Reactieve. Agressie. Pien S. Martens. Open Universiteit Heerlen
 REACTIEVE AGRESSIE Het Verband Tussen Negatieve Levensgebeurtenissen, 5-HTTLPR en Reactieve Agressie Pien S. Martens Open Universiteit Heerlen Naam student: Pien Sophie Martens Studentnummer: 850945172
REACTIEVE AGRESSIE Het Verband Tussen Negatieve Levensgebeurtenissen, 5-HTTLPR en Reactieve Agressie Pien S. Martens Open Universiteit Heerlen Naam student: Pien Sophie Martens Studentnummer: 850945172
3e Mirror meeting pren April :00 Session T, NVvA Symposium
 3e Mirror meeting pren 689 13 April 2017 14:00 Session T, NVvA Symposium steps since April 2016 The enquiry (June to August 2016) performed by the national bodies. Resulting in 550 comments. Three/Four
3e Mirror meeting pren 689 13 April 2017 14:00 Session T, NVvA Symposium steps since April 2016 The enquiry (June to August 2016) performed by the national bodies. Resulting in 550 comments. Three/Four
Persoonlijkheidskenmerken en cyberpesten onder jongeren van 11 tot 16 jaar:
 Persoonlijkheidskenmerken en cyberpesten onder jongeren van 11 tot 16 jaar: is er een relatie met een verkorte versie van de NVP-J? Personality Characteristics and Cyberbullying among youngsters of 11
Persoonlijkheidskenmerken en cyberpesten onder jongeren van 11 tot 16 jaar: is er een relatie met een verkorte versie van de NVP-J? Personality Characteristics and Cyberbullying among youngsters of 11
Classification of triangles
 Classification of triangles A triangle is a geometrical shape that is formed when 3 non-collinear points are joined. The joining line segments are the sides of the triangle. The angles in between the sides
Classification of triangles A triangle is a geometrical shape that is formed when 3 non-collinear points are joined. The joining line segments are the sides of the triangle. The angles in between the sides
Knelpunten in Zelfstandig Leren: Zelfregulerend leren, Stress en Uitstelgedrag bij HRM- Studenten van Avans Hogeschool s-hertogenbosch
 Knelpunten in Zelfstandig Leren: Zelfregulerend leren, Stress en Uitstelgedrag bij HRM- Studenten van Avans Hogeschool s-hertogenbosch Bottlenecks in Independent Learning: Self-Regulated Learning, Stress
Knelpunten in Zelfstandig Leren: Zelfregulerend leren, Stress en Uitstelgedrag bij HRM- Studenten van Avans Hogeschool s-hertogenbosch Bottlenecks in Independent Learning: Self-Regulated Learning, Stress
FOR DUTCH STUDENTS! ENGLISH VERSION NEXT PAGE
 FOR DUTCH STUDENTS! ENGLISH VERSION NEXT PAGE Tentamen Analyse 6 januari 203, duur 3 uur. Voeg aan het antwoord van een opgave altijd het bewijs, de berekening of de argumentatie toe. Als je een onderdeel
FOR DUTCH STUDENTS! ENGLISH VERSION NEXT PAGE Tentamen Analyse 6 januari 203, duur 3 uur. Voeg aan het antwoord van een opgave altijd het bewijs, de berekening of de argumentatie toe. Als je een onderdeel
Aim of this presentation. Give inside information about our commercial comparison website and our role in the Dutch and Spanish energy market
 Aim of this presentation Give inside information about our commercial comparison website and our role in the Dutch and Spanish energy market Energieleveranciers.nl (Energysuppliers.nl) Founded in 2004
Aim of this presentation Give inside information about our commercial comparison website and our role in the Dutch and Spanish energy market Energieleveranciers.nl (Energysuppliers.nl) Founded in 2004
SUPPLEMENTARY FIGURES AND TABLES
 Altered RECQL5 expression in urothelial bladder carcinoma increases cellular proliferation and makes RECQL5 helicase activity a novel target for chemotherapy SUPPLEMENTARY FIGURES AND TABLES Supplementary
Altered RECQL5 expression in urothelial bladder carcinoma increases cellular proliferation and makes RECQL5 helicase activity a novel target for chemotherapy SUPPLEMENTARY FIGURES AND TABLES Supplementary
De Relatie tussen Dagelijkse Stress, Negatief Affect en de Invloed van Bewegen
 De Relatie tussen Dagelijkse Stress, Negatief Affect en de Invloed van Bewegen The Association between Daily Hassles, Negative Affect and the Influence of Physical Activity Petra van Straaten Eerste begeleider
De Relatie tussen Dagelijkse Stress, Negatief Affect en de Invloed van Bewegen The Association between Daily Hassles, Negative Affect and the Influence of Physical Activity Petra van Straaten Eerste begeleider
Laboratory report. Independent testing of material surfaces. Analysis of leaching substances in treated wood samples conform guide line EU 10/2011
 Independent testing of material surfaces Laboratory report Analysis of leaching substances in treated wood samples conform guide line EU 10/2011 Customer Wasziederij De Vesting BV Trasweg 12 5712 BB Someren-Eind
Independent testing of material surfaces Laboratory report Analysis of leaching substances in treated wood samples conform guide line EU 10/2011 Customer Wasziederij De Vesting BV Trasweg 12 5712 BB Someren-Eind
Running head: OPVOEDSTIJL, EXTERNALISEREND PROLEEMGEDRAG EN ZELFBEELD
 1 Opvoedstijl en Externaliserend Probleemgedrag en de Mediërende Rol van het Zelfbeeld bij Dak- en Thuisloze Jongeren in Utrecht Parenting Style and Externalizing Problem Behaviour and the Mediational
1 Opvoedstijl en Externaliserend Probleemgedrag en de Mediërende Rol van het Zelfbeeld bij Dak- en Thuisloze Jongeren in Utrecht Parenting Style and Externalizing Problem Behaviour and the Mediational
Sedimentatie in Harderwijker Bocht ten gevolge van de strekdam bij Strand Horst Noord
 Sedimentatie in Harderwijker Bocht ten gevolge van de strekdam bij Strand Horst Noord In het gebied tussen de strekdammen bij Strand Horst Noord en de bebouwing van Harderwijk ligt een klein natuurgebied
Sedimentatie in Harderwijker Bocht ten gevolge van de strekdam bij Strand Horst Noord In het gebied tussen de strekdammen bij Strand Horst Noord en de bebouwing van Harderwijk ligt een klein natuurgebied
GOAL-STRIVING REASONS, PERSOONLIJKHEID EN BURN-OUT 1. Het effect van Goal-striving Reasons en Persoonlijkheid op facetten van Burn-out
 GOAL-STRIVING REASONS, PERSOONLIJKHEID EN BURN-OUT 1 Het effect van Goal-striving Reasons en Persoonlijkheid op facetten van Burn-out The effect of Goal-striving Reasons and Personality on facets of Burn-out
GOAL-STRIVING REASONS, PERSOONLIJKHEID EN BURN-OUT 1 Het effect van Goal-striving Reasons en Persoonlijkheid op facetten van Burn-out The effect of Goal-striving Reasons and Personality on facets of Burn-out
De Samenhang tussen Dagelijkse Stress, Emotionele Intimiteit en Affect bij Partners met een. Vaste Relatie
 De Samenhang tussen Dagelijkse Stress, Emotionele Intimiteit en Affect bij Partners met een Vaste Relatie The Association between Daily Stress, Emotional Intimacy and Affect with Partners in a Commited
De Samenhang tussen Dagelijkse Stress, Emotionele Intimiteit en Affect bij Partners met een Vaste Relatie The Association between Daily Stress, Emotional Intimacy and Affect with Partners in a Commited
De invloed van veerkracht op de relatie tussen pijn en psychische klachten bij revalidatiecliënten in een verpleeghuis.
 De invloed van veerkracht op de relatie tussen pijn en psychische klachten bij revalidatiecliënten in een verpleeghuis. The influence of resilience on the relationship between pain and psychological symptoms
De invloed van veerkracht op de relatie tussen pijn en psychische klachten bij revalidatiecliënten in een verpleeghuis. The influence of resilience on the relationship between pain and psychological symptoms
Academisch schrijven Inleiding
 - In this essay/paper/thesis I shall examine/investigate/evaluate/analyze Algemene inleiding van het werkstuk In this essay/paper/thesis I shall examine/investigate/evaluate/analyze To answer this question,
- In this essay/paper/thesis I shall examine/investigate/evaluate/analyze Algemene inleiding van het werkstuk In this essay/paper/thesis I shall examine/investigate/evaluate/analyze To answer this question,
Esther Lee-Varisco Matt Zhang
 Esther Lee-Varisco Matt Zhang Want to build a wine cellar Surface temperature varies daily, seasonally, and geologically Need reasonable depth to build the cellar for lessened temperature variations Building
Esther Lee-Varisco Matt Zhang Want to build a wine cellar Surface temperature varies daily, seasonally, and geologically Need reasonable depth to build the cellar for lessened temperature variations Building
De Relatie tussen Werkdruk, Pesten op het Werk, Gezondheidsklachten en Verzuim
 De Relatie tussen Werkdruk, Pesten op het Werk, Gezondheidsklachten en Verzuim The Relationship between Work Pressure, Mobbing at Work, Health Complaints and Absenteeism Agnes van der Schuur Eerste begeleider:
De Relatie tussen Werkdruk, Pesten op het Werk, Gezondheidsklachten en Verzuim The Relationship between Work Pressure, Mobbing at Work, Health Complaints and Absenteeism Agnes van der Schuur Eerste begeleider:
TE KOOP BOUWGROND N201 VERMOGENWEG MIJDRECHT COLLIERS INTERNATIONAL REAL ESTATE B.V. TE KOOP BOUWGROND N201 VERMOGENWEG MIJDRECHT
 TE KOOP BOUWGROND N201 VERMOGENWEG MIJDRECHT COLLIERS INTERNATIONAL REAL ESTATE B.V. TE KOOP BOUWGROND N201 VERMOGENWEG MIJDRECHT COLLIERS INTERNATIONAL REAL ESTATE B.V. TE KOOP/ TE HUUR ADRES - PLAATS
TE KOOP BOUWGROND N201 VERMOGENWEG MIJDRECHT COLLIERS INTERNATIONAL REAL ESTATE B.V. TE KOOP BOUWGROND N201 VERMOGENWEG MIJDRECHT COLLIERS INTERNATIONAL REAL ESTATE B.V. TE KOOP/ TE HUUR ADRES - PLAATS
Positieve, Negatieve en Depressieve Subklinische Psychotische Symptomen en het Effect van Stress en Sekse op deze Subklinische Psychotische Symptomen
 Positieve, Negatieve en Depressieve Subklinische Psychotische Symptomen en het Effect van Stress en Sekse op deze Subklinische Psychotische Symptomen Positive, Negative and Depressive Subclinical Psychotic
Positieve, Negatieve en Depressieve Subklinische Psychotische Symptomen en het Effect van Stress en Sekse op deze Subklinische Psychotische Symptomen Positive, Negative and Depressive Subclinical Psychotic
Denken is Doen? De cognitieve representatie van ziekte als determinant van. zelfmanagementgedrag bij Nederlandse, Turkse en Marokkaanse patiënten
 Denken is Doen? De cognitieve representatie van ziekte als determinant van zelfmanagementgedrag bij Nederlandse, Turkse en Marokkaanse patiënten met diabetes mellitus type 2 in de huisartsenpraktijk Thinking
Denken is Doen? De cognitieve representatie van ziekte als determinant van zelfmanagementgedrag bij Nederlandse, Turkse en Marokkaanse patiënten met diabetes mellitus type 2 in de huisartsenpraktijk Thinking
Macrofaunagemeenschap in het vooroeversuppletiegebied Ameland Midden tijdens de zomer 2009 (T 0 meting).
.") Macrofaunagemeenschap in het vooroeversuppletiegebied Ameland Midden tijdens de zomer 2009 (T 0 meting). Jeroen Wijsman 1, Kees Goudswaard 1, Erik Meesters 1 & Thomas Vanagt 2 Rapport C119/09 1 Wageningen
Macrofaunagemeenschap in het vooroeversuppletiegebied Ameland Midden tijdens de zomer 2009 (T 0 meting). Jeroen Wijsman 1, Kees Goudswaard 1, Erik Meesters 1 & Thomas Vanagt 2 Rapport C119/09 1 Wageningen
FOR DUTCH STUDENTS! ENGLISH VERSION NEXT PAGE. Toets Inleiding Kansrekening 1 7 februari 2011
 FOR DUTCH STUDENTS! ENGLISH VERSION NEXT PAGE Toets Inleiding Kansrekening 1 7 februari 2011 Voeg aan het antwoord van een opgave altijd het bewijs, de berekening of de argumentatie toe. Als je een onderdeel
FOR DUTCH STUDENTS! ENGLISH VERSION NEXT PAGE Toets Inleiding Kansrekening 1 7 februari 2011 Voeg aan het antwoord van een opgave altijd het bewijs, de berekening of de argumentatie toe. Als je een onderdeel
De Rol van Zelfregulatie, Motivatie en Eigen Effectiviteitsverwachting op het Volhouden
 De Rol van Zelfregulatie, Motivatie en Eigen Effectiviteitsverwachting op het Volhouden van Sporten en de Invloed van Egodepletie, Gewoonte en Geslacht The Role of Selfregulation, Motivation and Self-efficacy
De Rol van Zelfregulatie, Motivatie en Eigen Effectiviteitsverwachting op het Volhouden van Sporten en de Invloed van Egodepletie, Gewoonte en Geslacht The Role of Selfregulation, Motivation and Self-efficacy
Kwaliteit van Leven en Depressieve Symptomen van Mensen met Multiple Sclerose: De Modererende Invloed van Coping en Doelaanpassing
 Kwaliteit van Leven en Depressieve Symptomen van Mensen met Multiple Sclerose: De Modererende Invloed van Coping en Doelaanpassing Quality of Life and Depressive Symptoms of People with Multiple Sclerosis:
Kwaliteit van Leven en Depressieve Symptomen van Mensen met Multiple Sclerose: De Modererende Invloed van Coping en Doelaanpassing Quality of Life and Depressive Symptoms of People with Multiple Sclerosis:
Invloed van Mindfulness Training op Ouderlijke Stress, Emotionele Self-Efficacy. Beliefs, Aandacht en Bewustzijn bij Moeders
 Invloed van Mindfulness Training op Ouderlijke Stress, Emotionele Self-Efficacy Beliefs, Aandacht en Bewustzijn bij Moeders Influence of Mindfulness Training on Parental Stress, Emotional Self-Efficacy
Invloed van Mindfulness Training op Ouderlijke Stress, Emotionele Self-Efficacy Beliefs, Aandacht en Bewustzijn bij Moeders Influence of Mindfulness Training on Parental Stress, Emotional Self-Efficacy
Fysieke Activiteit bij 50-plussers. The Relationship between Self-efficacy, Intrinsic Motivation and. Physical Activity among Adults Aged over 50
 De relatie tussen eigen-effectiviteit 1 De Relatie tussen Eigen-effectiviteit, Intrinsieke Motivatie en Fysieke Activiteit bij 50-plussers The Relationship between Self-efficacy, Intrinsic Motivation and
De relatie tussen eigen-effectiviteit 1 De Relatie tussen Eigen-effectiviteit, Intrinsieke Motivatie en Fysieke Activiteit bij 50-plussers The Relationship between Self-efficacy, Intrinsic Motivation and
SAMPLE 11 = + 11 = + + Exploring Combinations of Ten + + = = + + = + = = + = = 11. Step Up. Step Ahead
 7.1 Exploring Combinations of Ten Look at these cubes. 2. Color some of the cubes to make three parts. Then write a matching sentence. 10 What addition sentence matches the picture? How else could you
7.1 Exploring Combinations of Ten Look at these cubes. 2. Color some of the cubes to make three parts. Then write a matching sentence. 10 What addition sentence matches the picture? How else could you
de Rol van Persoonlijkheid Eating: the Role of Personality
 De Relatie tussen Dagelijkse Stress en Emotioneel Eten: de Rol van Persoonlijkheid The Relationship between Daily Stress and Emotional Eating: the Role of Personality Arlette Nierich Open Universiteit
De Relatie tussen Dagelijkse Stress en Emotioneel Eten: de Rol van Persoonlijkheid The Relationship between Daily Stress and Emotional Eating: the Role of Personality Arlette Nierich Open Universiteit
Add the standing fingers to get the tens and multiply the closed fingers to get the units.
 Digit work Here's a useful system of finger reckoning from the Middle Ages. To multiply $6 \times 9$, hold up one finger to represent the difference between the five fingers on that hand and the first
Digit work Here's a useful system of finger reckoning from the Middle Ages. To multiply $6 \times 9$, hold up one finger to represent the difference between the five fingers on that hand and the first
Daylight saving time. Assignment
 Daylight saving time Daylight saving time (DST or summertime) is the arrangement by which clocks are advanced by one hour in spring and moved back in autumn to make the most of seasonal daylight Spring:
Daylight saving time Daylight saving time (DST or summertime) is the arrangement by which clocks are advanced by one hour in spring and moved back in autumn to make the most of seasonal daylight Spring:
De relatie tussen Stress Negatief Affect en Opvoedstijl. The relationship between Stress Negative Affect and Parenting Style
 De relatie tussen Stress Negatief Affect en Opvoedstijl The relationship between Stress Negative Affect and Parenting Style Jenny Thielman 1 e begeleider: mw. dr. Esther Bakker 2 e begeleider: mw. dr.
De relatie tussen Stress Negatief Affect en Opvoedstijl The relationship between Stress Negative Affect and Parenting Style Jenny Thielman 1 e begeleider: mw. dr. Esther Bakker 2 e begeleider: mw. dr.